

Review
doi: 10.1038/nrm2636.
Epub 2009 Feb 11.
Epidermal homeostasis: a balancing act of stem cells in the skin
Affiliations
- PMID: 19209183
- PMCID: PMC2760218
- DOI: 10.1038/nrm2636
Item in Clipboard
Review
Epidermal homeostasis: a balancing act of stem cells in the skin
Nat Rev Mol Cell Biol.
2009 Mar.
Abstract
The skin epidermis and its array of appendages undergo ongoing renewal by a process called homeostasis. Stem cells in the epidermis have a crucial role in maintaining tissue homeostasis by providing new cells to replace those that are constantly lost during tissue turnover or following injury. Different resident skin stem cell pools contribute to the maintenance and repair of the various epidermal tissues of the skin, including interfollicular epidermis, hair follicles and sebaceous glands. Interestingly, the basic mechanisms and signalling pathways that orchestrate epithelial morphogenesis in the skin are reused during adult life to regulate skin homeostasis.
Figures
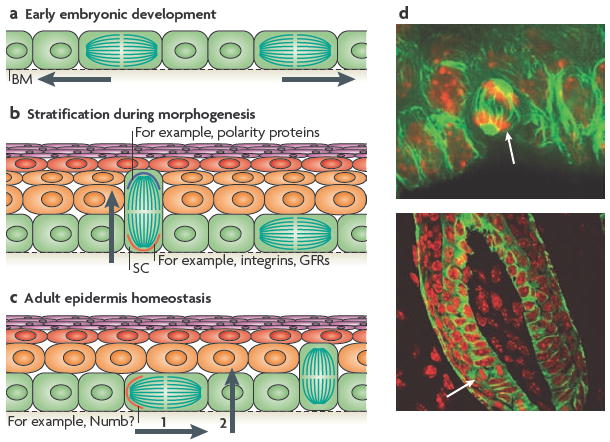
a During the early stages of embryonic skin development, most cell divisions are symmetric and parallel to the basement membrane (BM), which ensures the growth of the surface of the developing embryo and maintains the epithelium as a single layer. b During epidermal stratification, ~70% of the cell divisions become asymmetric, such that the mitotic spindle is perpendicular to the basement membrane. This allows the development of suprabasal cells that terminally differentiate and establish the skin barrier (indicated by the arrow). In this mode of asymmetric cell division, different cell fate determinants are segregated unequally between the two daughter cells. The basal cell segregates the integrins and growth factor receptors (GFRs), which provide survival and proliferative cues to the stem cell (SC). Polarity proteins are concentrated to the apical surface and presumably become preferentially distributed to the suprabasal cell. c During epidermal homeostasis in the adult tail skin, asymmetric cell divisions occur with the plane of division parallel to the BM, such that only one daughter cell inherits a cell fate determinant, such as Numb, and remains a SC (step 1), whereas the other becomes committed to terminal differentiation, and probably undergoes delamination to reach the suprabasal layers (step 2). d Examples of asymmetric spindle orientations (white arrow) in the oral epithelium, which are similar to those seen in the embryonic epidermis (upper panel) and in the adult hair bulb (lower panel). Nuclei are shown in red and microtubules in green. Images in part d are courtesy of T. Lechler and E.F., Rockefeller University, New York, USA.
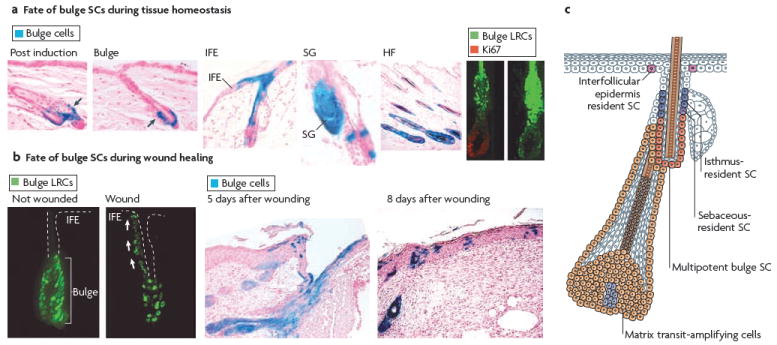
a During telogen, bulge stem cells (SCs) are specifically marked by β-galactosidase (LacZ; blue) by inducing the activity of an inducible Cre recombinase expressed under the control of a bulge promoter. When hair follicles (HFs) undergo cycling, all cells of newly formed HFs are marked, demonstrating that bulge SCs fuel normal follicle homeostasis. Bulge SCs can also contribute to the formation of interfollicular epidermis (IFE) and sebaceous glands (SGs), although this phenomenon is rare and, in most cases, IFE cells are not derived from bulge cells. This demonstrates that the IFE can be maintained independently of bulge SCs during tissue homeostasis. The bulge SCs, which are shown as green histone H2B–green fluorescent protein-labelled retaining cells (LRCs), exit the SC niche and actively proliferate (as shown here by Ki67 immunoreactivity) to provide cells that initiate HF regeneration. b Following wounding, the bulge SCs, become activated and migrate upward to repair the IFE. c Schematic representation of the skin epidermis with the different resident SC compartments and transit-amplifying progeny identified. Bulge SCs are multipotent, residing in the permanent portion of the HF. IFE SCs reside in the basal layer of the epidermis. Resident progenitors of the isthmus and SG reside in the outer root sheath that is above the bulge and below the SG. It is not clear whether these two resident progenitors are equivalent. Figure part a is reproduced, with permission, from REF. © (2004) American Association for the Advancement of Science, and from Nature Biotechnology REF. © (2004) Macmillan Publishers Ltd. All rights reserved. Figure part b, is reproduced, with permission, from REF. © (2004) American Association for the Advancement of Science, and from Nature Medicine REF. © (2005) Macmillan Publishers Ltd. All rights reserved.
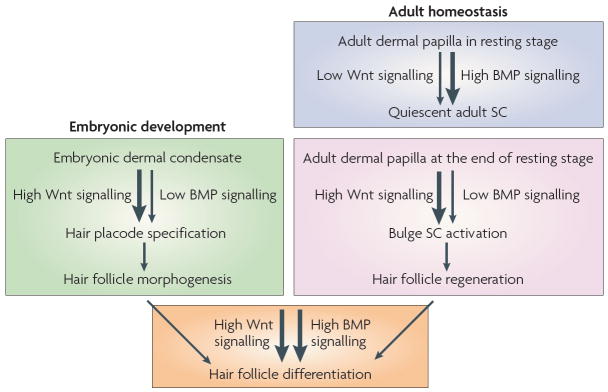
Loss- and gain-of-function studies in mice highlight the different functions of Wnt/β-catenin and bone morphogenetic protein (BMP) signalling during morphogenesis and adult skin homeostasis. During hair follicle development, Wnt/β-catenin signalling is required to specify the hair follicle fate in the undifferentiated basal epidermis. During the adult hair cycle, increased Wnt signalling promotes stem cell (SC) activation to initiate the growth of a new hair during the transition from resting to growing stage. An even more robust response to Wnt signalling is involved later as matrix cells specifically commit to terminal differentiation along the hair shaft lineage. BMP signals are transmitted from the underlying mesenchyme to the epidermis. As dermal condensates form, the prospective dermal papilla expresses the BMP inhibitor noggin. Noggin is required for normal follicle development and stimulates lymphoid enhancer-binding factor 1 (LEF1) expression and Wnt signalling. During the adult hair cycle, active BMP signalling propels bulge SC quiescence during the resting stage. During the transition from resting to growing stage of the hair cycle, the levels of dermal BMPs and BMP inhibitors change, resulting in a net inhibition of BMP signalling and SC activation. As the new follicle matures, activation of BMP receptor signalling is essential for the differentiation of transit-amplifying matrix cells to form the hair shaft and its channel.
References
-
- Koster MI, Roop DR. Mechanisms regulating epithelial stratification. Annu Rev Cell Dev Biol. 2007;23:93–113. - PubMed
-
- Fuchs E, Green H. Changes in keratin gene expression during terminal differentiation of the keratinocyte. Cell. 1980;19:1033–1042. - PubMed
-
- Candi E, Schmidt R, Melino G. The cornified envelope: a model of cell death in the skin. Nature Rev Mol Cell Biol. 2005;6:328–340. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
