

Evidence for bidentate substrate binding as the basis for the K48 linkage specificity of otubain 1
- PMID: 19211026
- PMCID: PMC2682458
- DOI: 10.1016/j.jmb.2008.12.085
Evidence for bidentate substrate binding as the basis for the K48 linkage specificity of otubain 1
Abstract
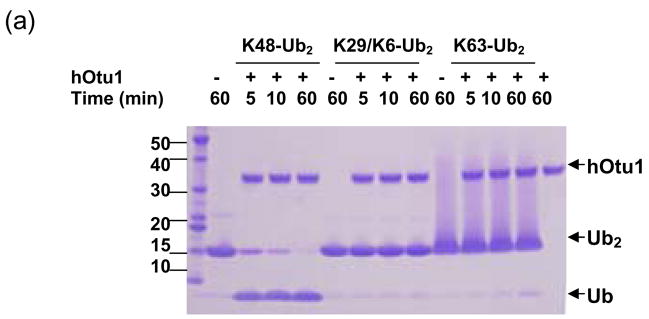
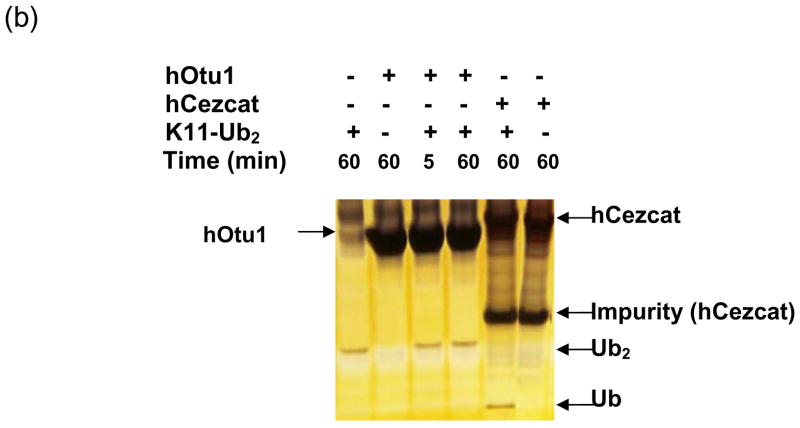
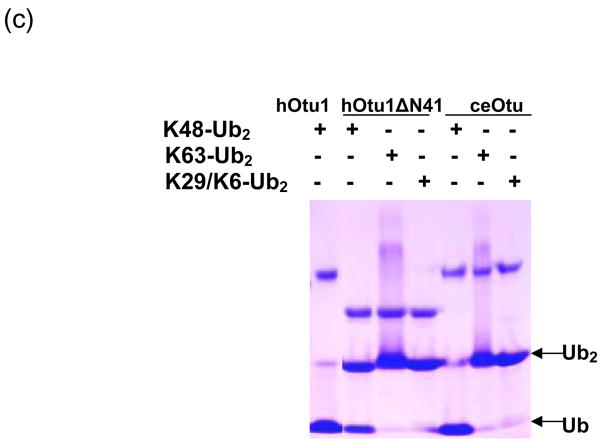
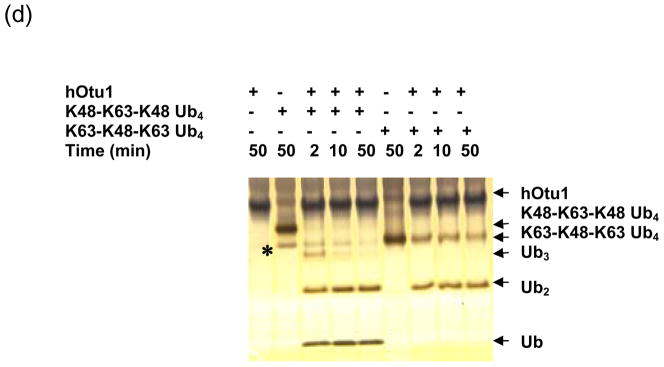
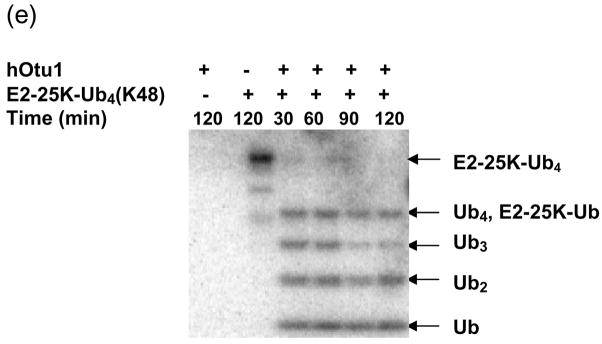
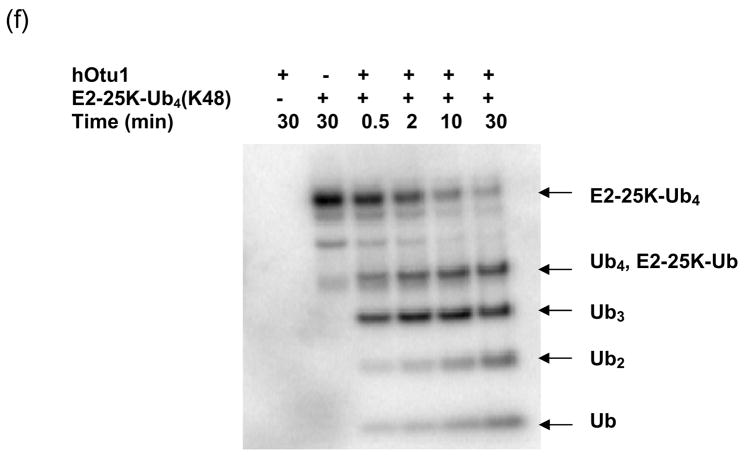
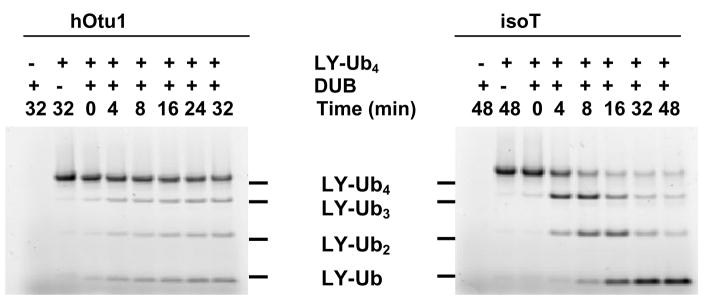
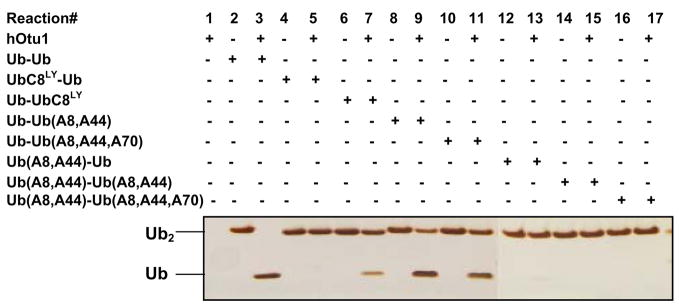
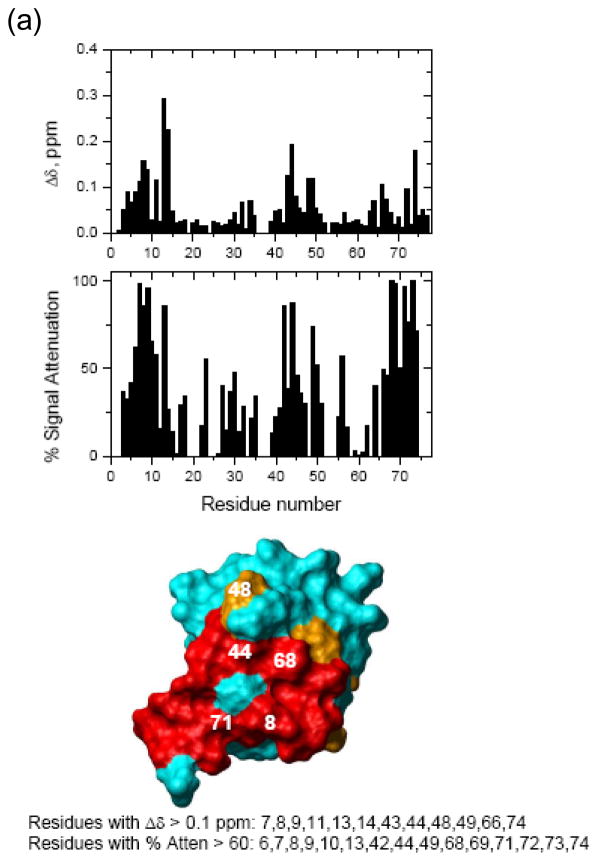
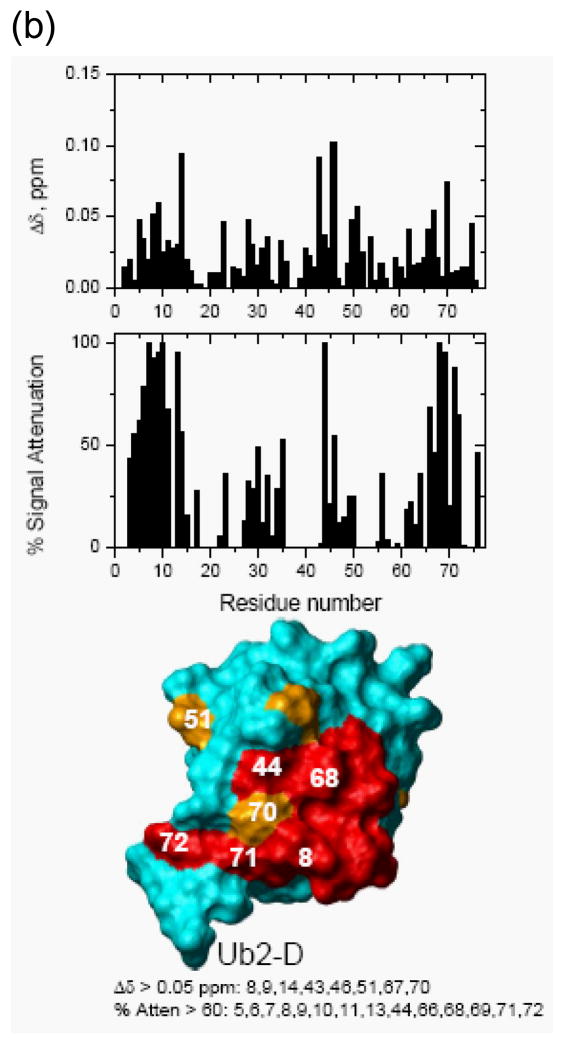
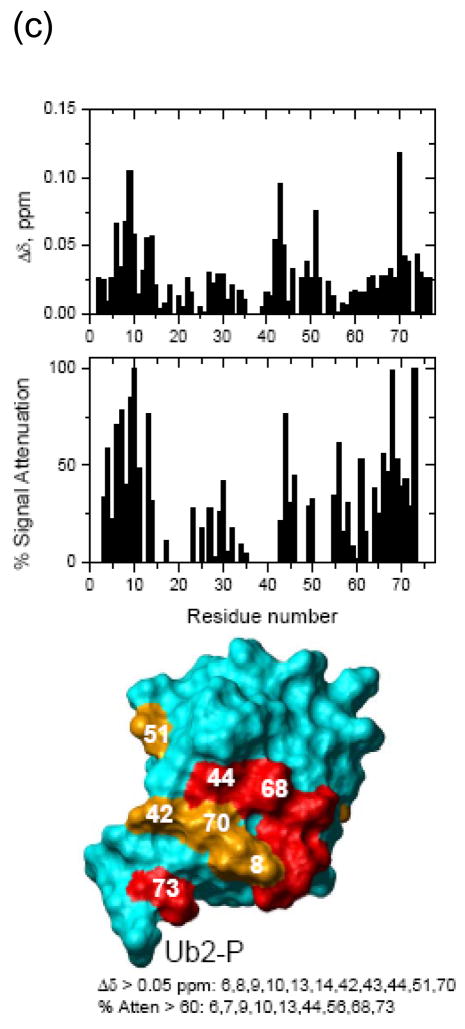
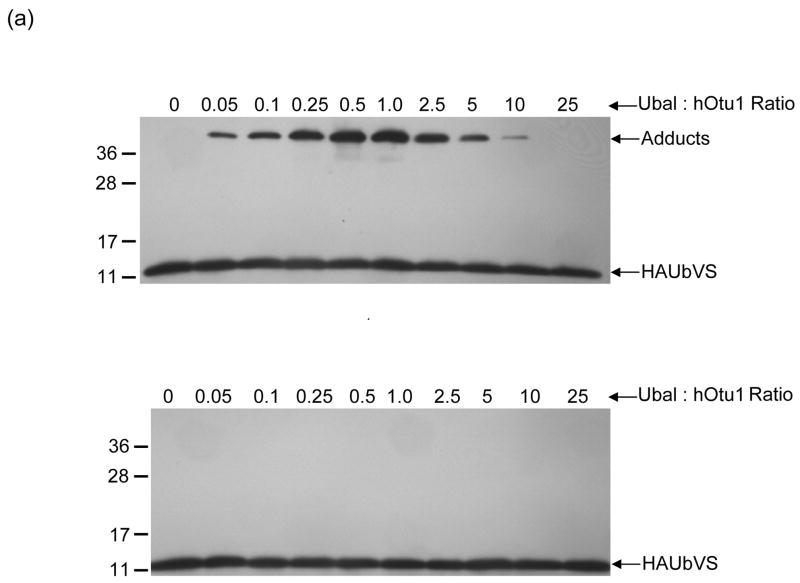
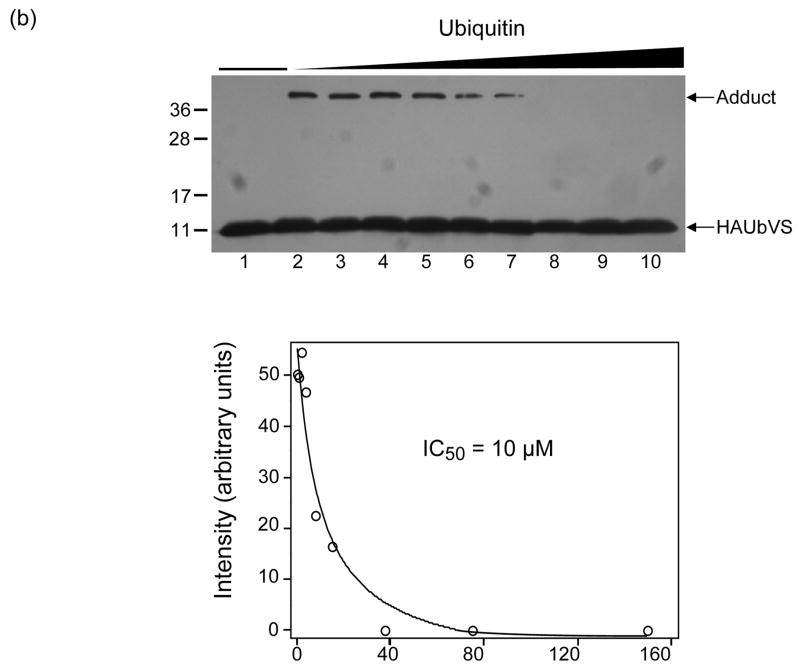
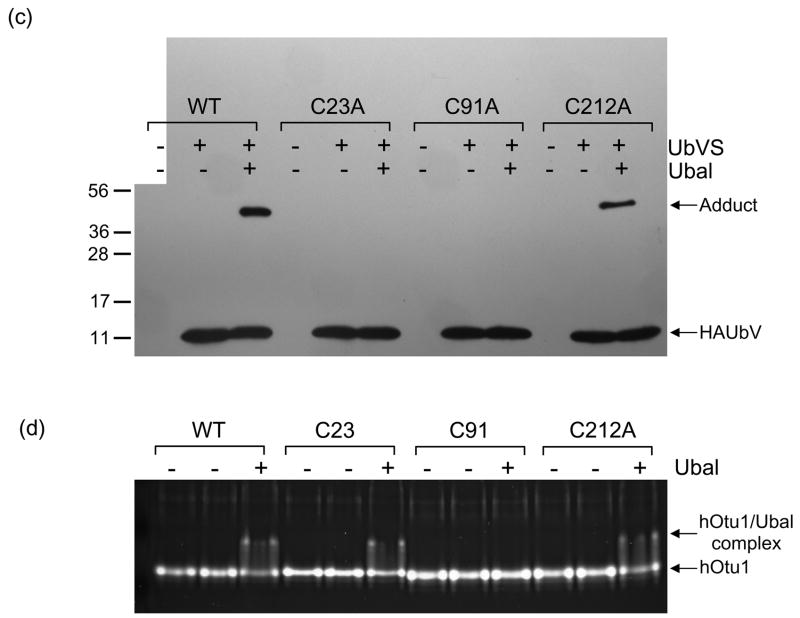
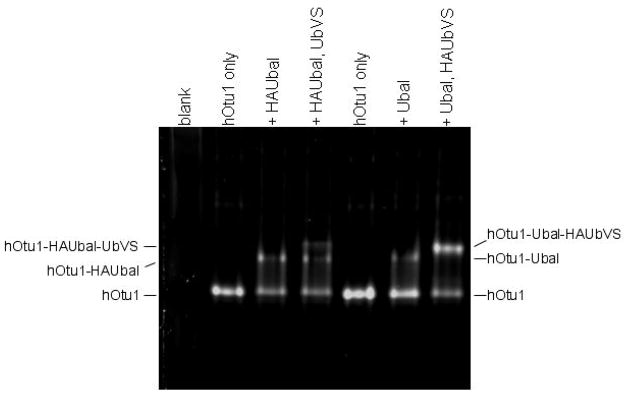
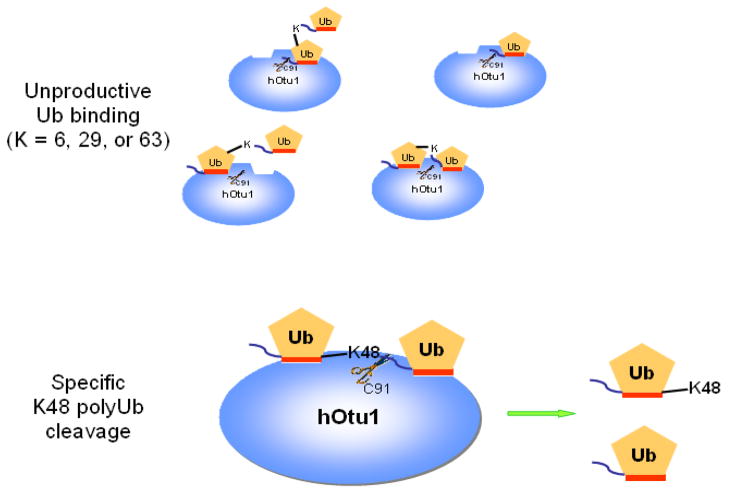
Otubain 1 belongs to the ovarian tumor (OTU) domain class of cysteine protease deubiquitinating enzymes. We show here that human otubain 1 (hOtu1) is highly linkage-specific, cleaving Lys48 (K48)-linked polyubiquitin but not K63-, K29-, K6-, or K11-linked polyubiquitin, or linear alpha-linked polyubiquitin. Cleavage is not limited to either end of a polyubiquitin chain, and both free and substrate-linked polyubiquitin are disassembled. Intriguingly, cleavage of K48-diubiquitin by hOtu1 can be inhibited by diubiquitins of various linkage types, as well as by monoubiquitin. NMR studies and activity assays suggest that both the proximal and distal units of K48-diubiquitin bind to hOtu1. Reaction of Cys23 with ubiquitin-vinylsulfone identified a ubiquitin binding site that is distinct from the active site, which includes Cys91. Occupancy of the active site is needed to enable tight binding to the second site. We propose that distinct binding sites for the ubiquitins on either side of the scissile bond allow hOtu1 to discriminate among different isopeptide linkages in polyubiquitin substrates. Bidentate binding may be a general strategy used to achieve linkage-specific deubiquitination.
Figures
References
-
- Hochstrasser M. Lingering mysteries of ubiquitin-chain assembly. Cell. 2006;124:27–34. - PubMed
-
- Pickart CM. Ubiquitin enters the new millennium. Mol Cell. 2001;8:499–504. - PubMed
-
- Pickart CM, Eddins MJ. Ubiquitin: structures, functions, mechanisms. Biochim Biophys Acta. 2004;1695:55–72. - PubMed
-
- Peng J, Schwartz D, Elias JE, Thoreen CC, Cheng D, Marsischky G, Roelofs J, Finley D, Gygi SP. A proteomics approach to understanding protein ubiquitination. Nat Biotechnol. 2003;21:921–6. - PubMed
-
- Pickart CM, Fushman D. Polyubiquitin chains: polymeric protein signals. Curr Opin Chem Biol. 2004;8:610–6. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
