

Alternative splicing of anciently exonized 5S rRNA regulates plant transcription factor TFIIIA
- PMID: 19211543
- PMCID: PMC2675980
- DOI: 10.1101/gr.086876.108
Alternative splicing of anciently exonized 5S rRNA regulates plant transcription factor TFIIIA
Abstract
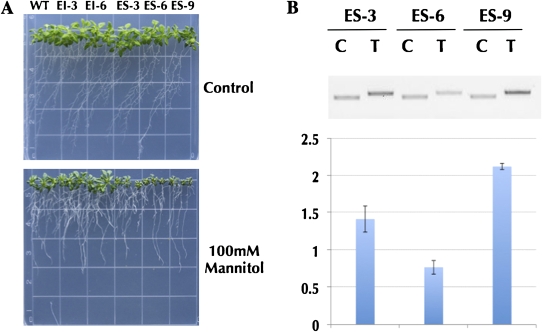
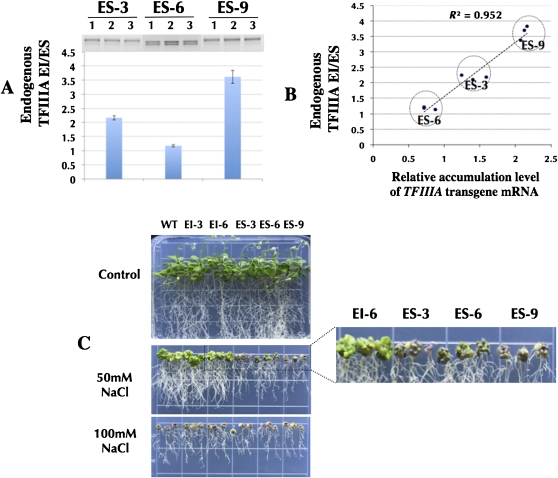
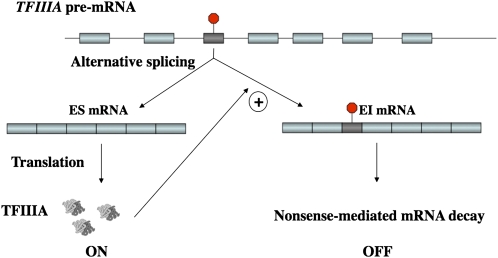
Identifying conserved alternative splicing (AS) events among evolutionarily distant species can prioritize AS events for functional characterization and help uncover relevant cis- and trans-regulatory factors. A genome-wide search for conserved cassette exon AS events in higher plants revealed the exonization of 5S ribosomal RNA (5S rRNA) within the gene of its own transcription regulator, TFIIIA (transcription factor for polymerase III A). The 5S rRNA-derived exon in TFIIIA gene exists in all representative land plant species but not in green algae and nonplant species, suggesting it is specific to land plants. TFIIIA is essential for RNA polymerase III-based transcription of 5S rRNA in eukaryotes. Integrating comparative genomics and molecular biology revealed that the conserved cassette exon derived from 5S rRNA is coupled with nonsense-mediated mRNA decay. Utilizing multiple independent Arabidopsis overexpressing TFIIIA transgenic lines under osmotic and salt stress, strong accordance between phenotypic and molecular evidence reveals the biological relevance of AS of the exonized 5S rRNA in quantitative autoregulation of TFIIIA homeostasis. Most significantly, this study provides the first evidence of ancient exaptation of 5S rRNA in plants, suggesting a novel gene regulation model mediated by the AS of an anciently exonized noncoding element.
Figures

References
-
- Barbazuk W.B., Fu Y., McGinnis K.M. Genome-wide analyses of alternative splicing in plants: Opportunities and challenges. Genome Res. 2008;18:1381–1392. - PubMed
-
- Beelman C.A., Parker R. Differential effects of translational inhibition in cis and in trans on the decay of the unstable yeast MFA2 mRNA. J. Biol. Chem. 1994;269:9687–9692. - PubMed
-
- Beitz E. TeXshade: Shading and labeling of multiple sequence alignments using LaTeX2ε. Bioinformatics. 2000;16:135–139. - PubMed
-
- Bejerano G., Lowe C.B., Ahituv N., King B., Siepel A., Salama S.R., Rubin E.M., Kent W.J., Haussler D. A distal enhancer and an ultraconserved exon are derived from a novel retroposon. Nature. 2006;441:87–90. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials