

Palmitoylation controls the catalytic activity and subcellular distribution of phosphatidylinositol 4-kinase II{alpha}
- PMID: 19211550
- PMCID: PMC2665123
- DOI: 10.1074/jbc.M900724200
Palmitoylation controls the catalytic activity and subcellular distribution of phosphatidylinositol 4-kinase II{alpha}
Abstract
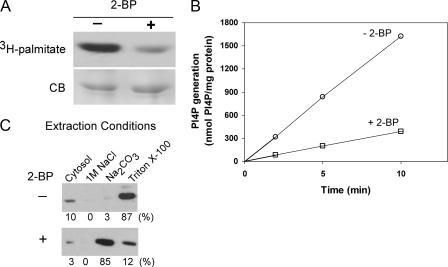
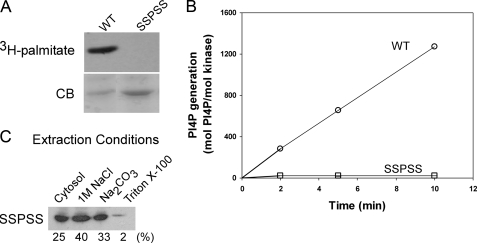
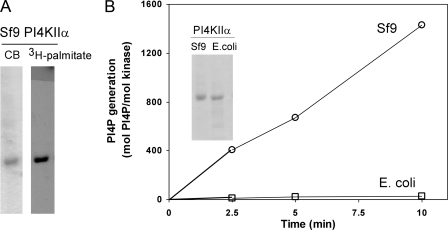
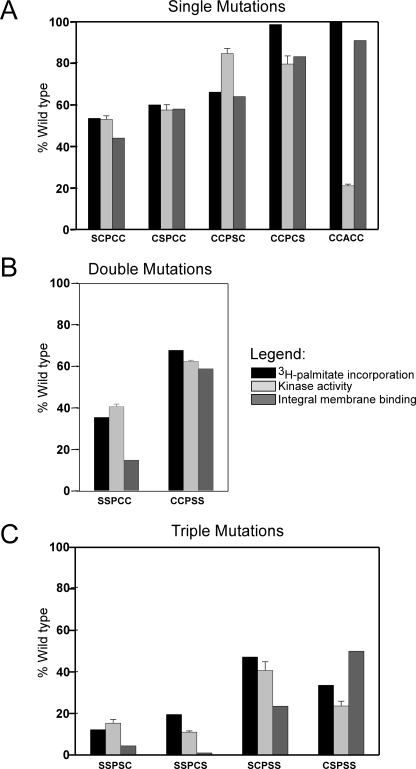
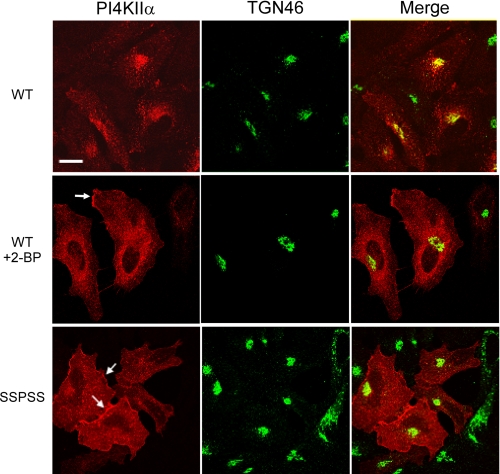
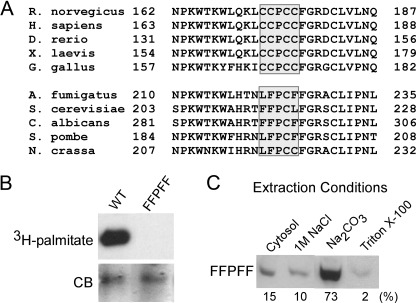
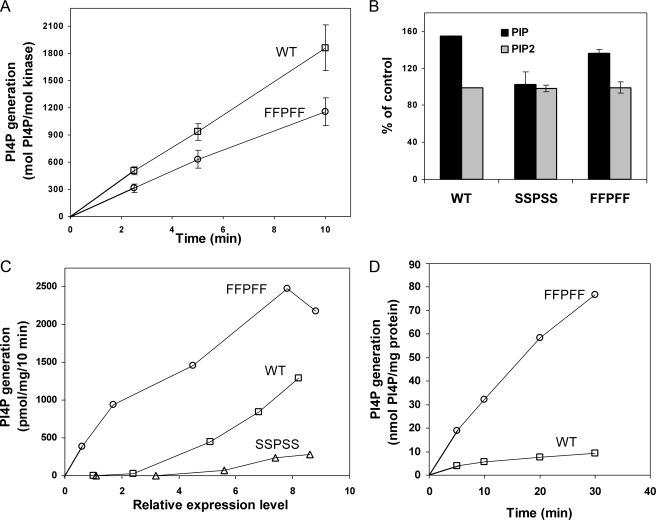
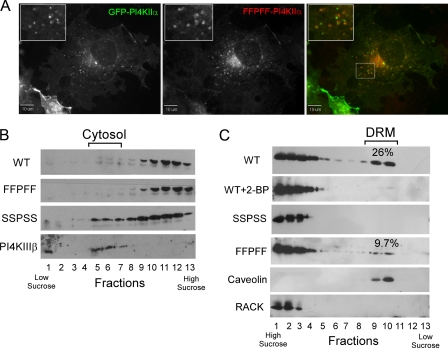
Phosphatidylinositol 4-kinases play essential roles in cell signaling and membrane trafficking. They are divided into type II and III families, which have distinct structural and enzymatic properties and are essentially unrelated in sequence. Mammalian cells express two type II isoforms, phosphatidylinositol 4-kinase IIalpha (PI4KIIalpha) and IIbeta (PI4KIIbeta). Nearly all of PI4KIIalpha, and about half of PI4KIIbeta, associates integrally with membranes, requiring detergent for solubilization. This tight membrane association is because of palmitoylation of a cysteine-rich motif, CCPCC, located within the catalytic domains of both type II isoforms. Deletion of this motif from PI4KIIalpha converts the kinase from an integral to a tightly bound peripheral membrane protein and abrogates its catalytic activity ( Barylko, B., Gerber, S. H., Binns, D. D., Grichine, N., Khvotchev, M., Sudhof, T. C., and Albanesi, J. P. (2001) J. Biol. Chem. 276, 7705-7708 ). Here we identify the first two cysteines in the CCPCC motif as the principal sites of palmitoylation under basal conditions, and we demonstrate the importance of the central proline for enzymatic activity, although not for membrane binding. We further show that palmitoylation is critical for targeting PI4KIIalpha to the trans-Golgi network and for enhancement of its association with low buoyant density membrane fractions, commonly termed lipid rafts. Replacement of the four cysteines in CCPCC with a hydrophobic residue, phenylalanine, substantially restores catalytic activity of PI4KIIalpha in vitro and in cells without restoring integral membrane binding. Although this FFPFF mutant displays a perinuclear distribution, it does not strongly co-localize with wild-type PI4KIIalpha and associates more weakly with lipid rafts.
Figures
References
-
- Di Paolo, G., and De Camilli, P. (2006) Nature 443651 -657 - PubMed
-
- De Matteis, M. A., and D'Angelo, G. (2007) Biochem. Soc. Symp. 74107 -116 - PubMed
-
- Fruman, D. A., Meyers, R. E., and Cantley, L. C. (1998) Annu. Rev. Biochem. 67 481-507 - PubMed
-
- Heilmeyer, L. M., Jr., Vereb, G., Jr., Vereb, G., Kakuk, A., and Szivak, I. (2003) IUBMB Life 55 59-65 - PubMed
-
- Balla, A., and Balla, T. (2006) Trends Cell Biol. 16351 -361 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
