

Sl-IAA3, a tomato Aux/IAA at the crossroads of auxin and ethylene signalling involved in differential growth
- PMID: 19213814
- PMCID: PMC2657550
- DOI: 10.1093/jxb/erp009
Sl-IAA3, a tomato Aux/IAA at the crossroads of auxin and ethylene signalling involved in differential growth
Abstract
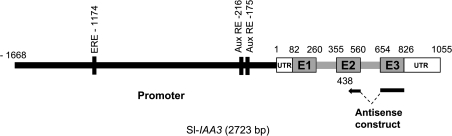
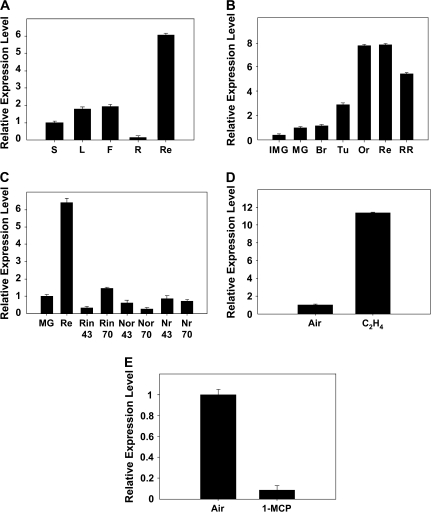
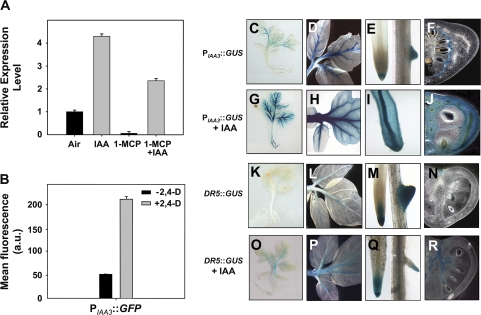
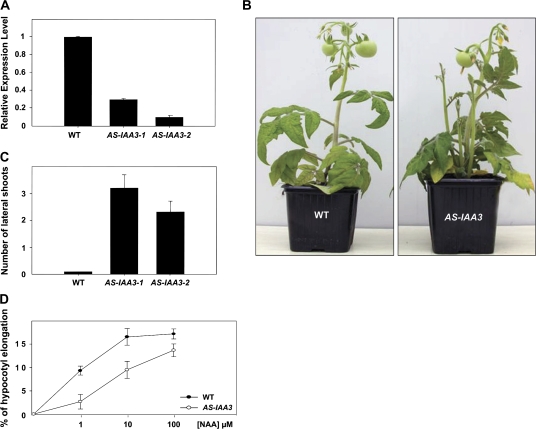
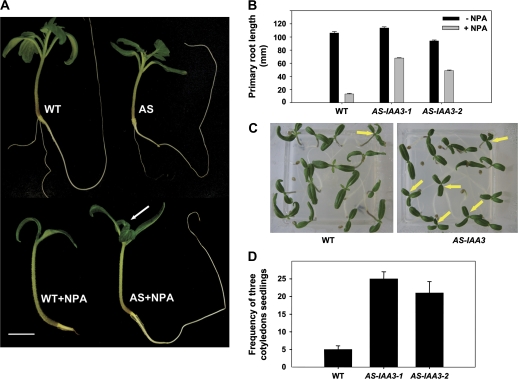
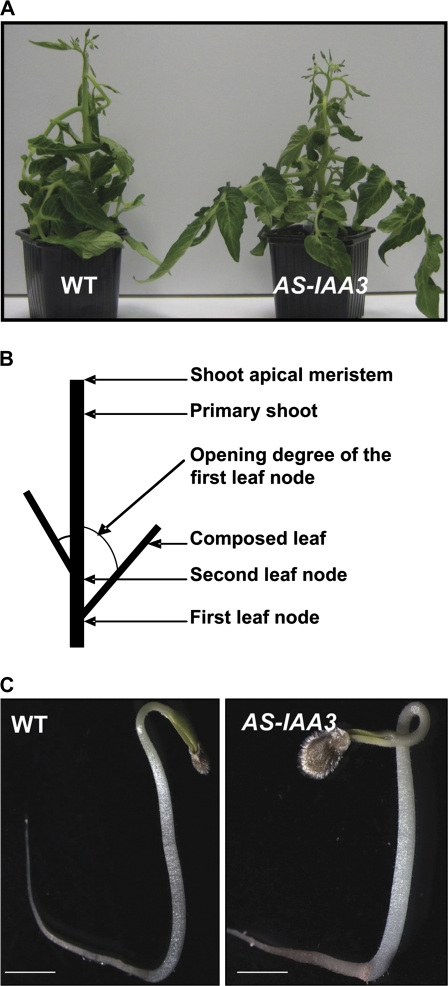
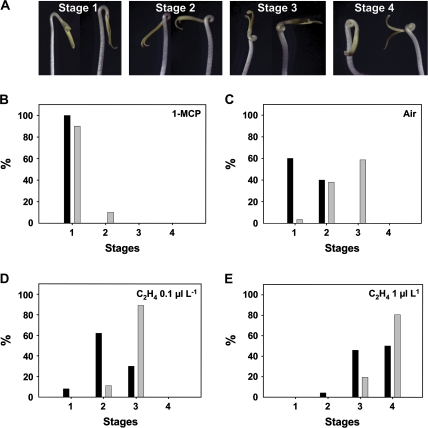
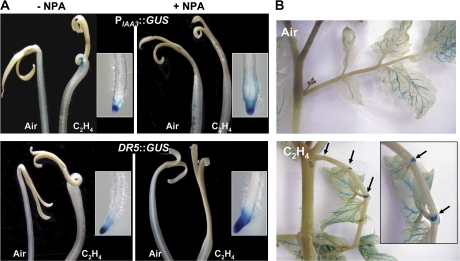
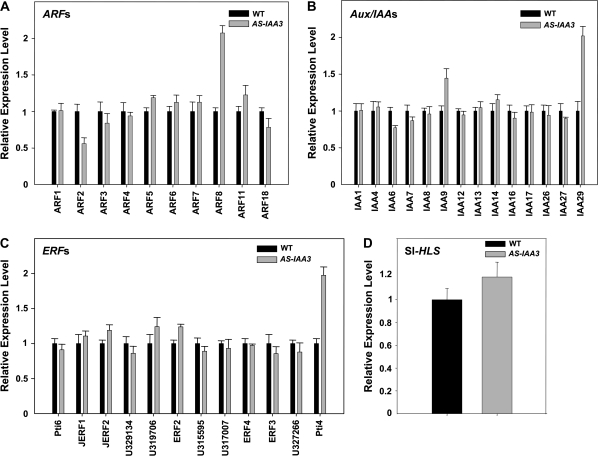
Whereas the interplay of multiple hormones is essential for most plant developmental processes, the key integrating molecular players remain largely undiscovered or uncharacterized. It is shown here that a member of the tomato auxin/indole-3-acetic acid (Aux/IAA) gene family, Sl-IAA3, intersects the auxin and ethylene signal transduction pathways. Aux/IAA genes encode short-lived transcriptional regulators central to the control of auxin responses. Their functions have been defined primarily by dominant, gain-of-function mutant alleles in Arabidopsis. The Sl-IAA3 gene encodes a nuclear-targeted protein that can repress transcription from auxin-responsive promoters. Sl-IAA3 expression is auxin and ethylene dependent, is regulated on a tight tissue-specific basis, and is associated with tissues undergoing differential growth such as in epinastic petioles and apical hook. Antisense down-regulation of Sl-IAA3 results in auxin and ethylene-related phenotypes, including altered apical dominance, lower auxin sensitivity, exaggerated apical hook curvature in the dark and reduced petiole epinasty in the light. The results provide novel insights into the roles of Aux/IAAs and position the Sl-IAA3 protein at the crossroads of auxin and ethylene signalling in tomato.
Figures
Comment in
-
Tomato Aux/IAA3 and HOOKLESS are important actors of the interplay between auxin and ethylene during apical hook formation.Plant Signal Behav. 2009 Jun;4(6):559-60. doi: 10.1093/jxb/erp009. Epub 2009 Jun 17. Plant Signal Behav. 2009. PMID: 19816148 Free PMC article.
References
-
- Abel S, Nguyen MD, Chow W, Theologis A. ACS4, a primary indole acetic acid-responsive gene encoding 1-aminocyclopropane-1-carboxylate synthase in Arabidopsis thaliana: structural, characterization, expression in Escherichia coli, and expression characteristics in response to auxin. Journal of Biological Chemistry. 1995;270:19093–19099. - PubMed
-
- Bleecker AB, Estelle MA, Somerville C, Kende H. Insensitivity to ethylene conferred by a dominant mutation in Arabidopsis thaliana. Science. 1988;26:1086–1089. - PubMed
-
- Chae HS, Cho YG, Park MY, Lee MC, Eun MY, Kang BG, Kim WT. Hormonal cross-talk between auxin and ethylene differentially regulates the expression of two members of the 1-aminocyclopropane-1-carboxylate oxidase gene family in rice (Oryza sativa L) Plant and Cell Physiology. 2000;41:354–362. - PubMed
-
- Dharmasiri N, Dharmasiri S, Estelle M. The F-box protein TIR1 is an auxin receptor. Nature. 2005a;435:441–445. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
