

ASK1 and ASK2 differentially regulate the counteracting roles of apoptosis and inflammation in tumorigenesis
- PMID: 19214184
- PMCID: PMC2670860
- DOI: 10.1038/emboj.2009.32
ASK1 and ASK2 differentially regulate the counteracting roles of apoptosis and inflammation in tumorigenesis
Abstract
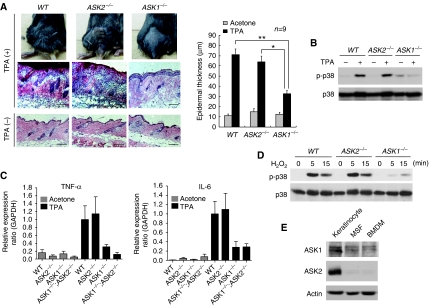
Apoptosis and inflammation generally exert opposite effects on tumorigenesis: apoptosis serves as a barrier to tumour initiation, whereas inflammation promotes tumorigenesis. Although both events are induced by various common stressors, relatively little is known about the stress-induced signalling pathways regulating these events in tumorigenesis. Here, we show that stress-activated MAP3Ks, ASK1 and ASK2, which are involved in cellular responses to various stressors such as reactive oxygen species, differentially regulate the initiation and promotion of tumorigenesis. ASK2 in cooperation with ASK1 functioned as a tumour suppressor by exerting proapoptotic activity in epithelial cells, which was consistent with the reduction in ASK2 expression in human cancer cells and tissues. In contrast, ASK1-dependent cytokine production in inflammatory cells promoted tumorigenesis. Our findings suggest that ASK1 and ASK2 are critically involved in tumorigenesis by differentially regulating apoptosis and inflammation.
Figures







References
-
- Azad N, Rojanasakul Y, Vallyathan V (2008) Inflammation and lung cancer: roles of reactive oxygen/nitrogen species. J Toxicol Environ Health B Crit Rev 11: 1–15 - PubMed
-
- Bode AM, Dong Z (2003) Mitogen-activated protein kinase activation in UV-induced signal transduction. Sci STKE 2003: RE2. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
