

Single nucleotide variants of the TGACTCA motif modulate energetics and orientation of binding of the Jun-Fos heterodimeric transcription factor
- PMID: 19215067
- PMCID: PMC2693225
- DOI: 10.1021/bi802068s
Single nucleotide variants of the TGACTCA motif modulate energetics and orientation of binding of the Jun-Fos heterodimeric transcription factor
Abstract
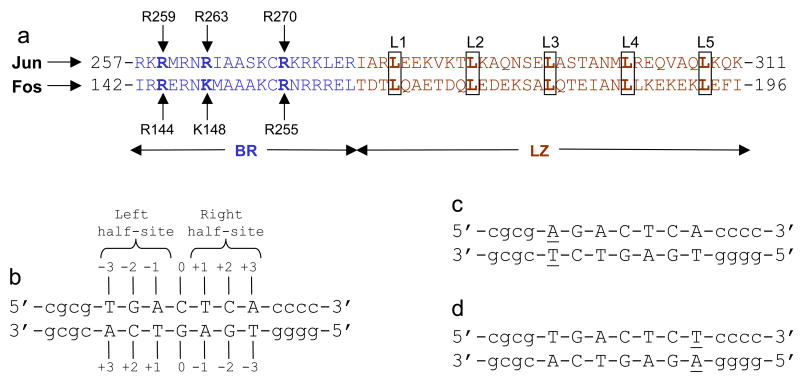
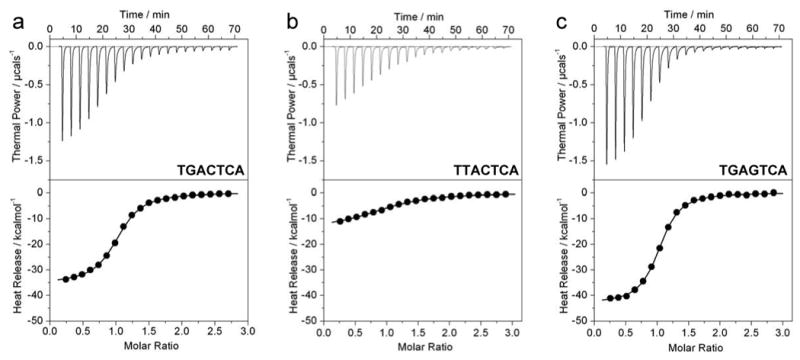
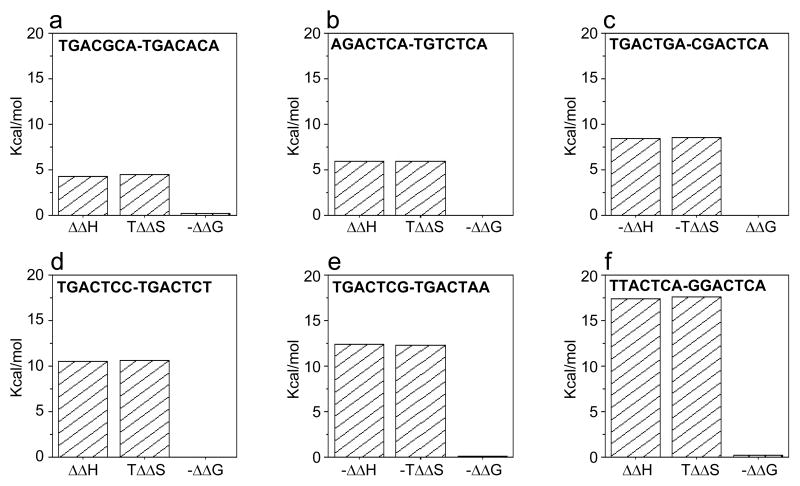
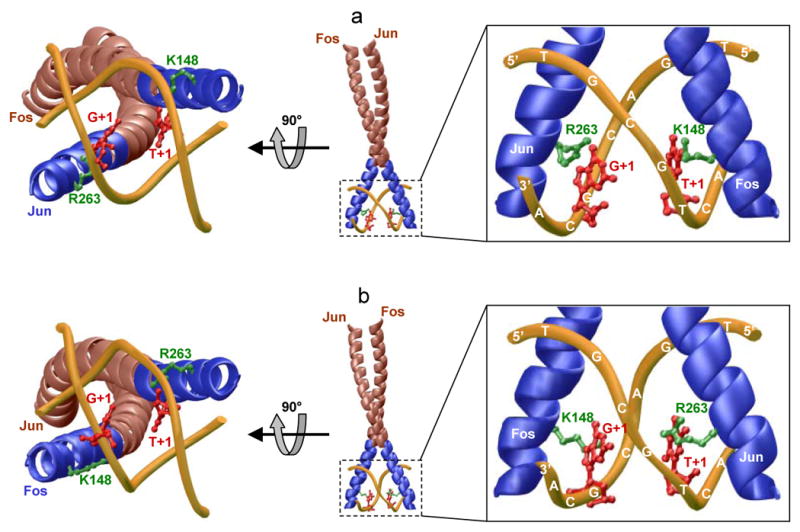
The Jun-Fos heterodimeric transcription factor is the terminal link between the transfer of extracellular information in the form of growth factors and cytokines to the site of DNA transcription within the nucleus in a wide variety of cellular processes central to health and disease. Here, using isothermal titration calorimetry, we report detailed thermodynamics of the binding of bZIP domains of Jun-Fos heterodimer to synthetic dsDNA oligos containing the TGACTCA cis element and all possible single nucleotide variants thereof encountered widely within the promoters of a diverse array of genes. Our data show that Jun-Fos heterodimer tolerates single nucleotide substitutions and binds to TGACTCA variants with affinities in the physiologically relevant micromolar to submicromolar range. The energetics of binding are richly favored by enthalpic forces and opposed by entropic changes across the entire spectrum of TGACTCA variants in agreement with the notion that protein-DNA interactions are largely driven by electrostatic interactions and intermolecular hydrogen bonding. Of particular interest is the observation that the Jun-Fos heterodimer binds to specific TGACTCA variants in a preferred orientation. Our 3D atomic models reveal that such orientational preference results from asymmetric binding and may in part be attributable to chemically distinct but structurally equivalent residues R263 and K148 located within the basic regions of Jun and Fos, respectively. Taken together, our data suggest that the single nucleotide variants of the TGACTCA motif modulate energetics and orientation of binding of the Jun-Fos heterodimer and that such behavior may be a critical determinant of differential regulation of specific genes under the control of this transcription factor. Our study also bears important consequences for the occurrence of single nucleotide polymorphisms within the TGACTCA cis element at specific gene promoters between different individuals.
Figures

Similar articles
-
DNA plasticity is a key determinant of the energetics of binding of Jun-Fos heterodimeric transcription factor to genetic variants of TGACGTCA motif.Biochemistry. 2009 Dec 29;48(51):12213-22. doi: 10.1021/bi901392k. Biochemistry. 2009. PMID: 19921846 Free PMC article.
-
Coupling of folding and DNA-binding in the bZIP domains of Jun-Fos heterodimeric transcription factor.Arch Biochem Biophys. 2008 May 1;473(1):48-60. doi: 10.1016/j.abb.2008.02.024. Epub 2008 Feb 26. Arch Biochem Biophys. 2008. PMID: 18316037
-
Long-range electrostatic interactions influence the orientation of Fos-Jun binding at AP-1 sites.J Mol Biol. 2001 Jan 19;305(3):411-27. doi: 10.1006/jmbi.2000.4286. J Mol Biol. 2001. PMID: 11152600
-
Close encounters of many kinds: Fos-Jun interactions that mediate transcription regulatory specificity.Oncogene. 2001 Apr 30;20(19):2438-52. doi: 10.1038/sj.onc.1204385. Oncogene. 2001. PMID: 11402339 Review.
-
Encounters with Fos and Jun on the road to AP-1.Semin Cancer Biol. 1990 Feb;1(1):19-26. Semin Cancer Biol. 1990. PMID: 2133107 Review.
Cited by
-
DNA plasticity is a key determinant of the energetics of binding of Jun-Fos heterodimeric transcription factor to genetic variants of TGACGTCA motif.Biochemistry. 2009 Dec 29;48(51):12213-22. doi: 10.1021/bi901392k. Biochemistry. 2009. PMID: 19921846 Free PMC article.
-
The N-Terminus of the Floral Arabidopsis TGA Transcription Factor PERIANTHIA Mediates Redox-Sensitive DNA-Binding.PLoS One. 2016 Apr 29;11(4):e0153810. doi: 10.1371/journal.pone.0153810. eCollection 2016. PLoS One. 2016. PMID: 27128442 Free PMC article.
-
Control of osteocyte dendrite formation by Sp7 and its target gene osteocrin.Nat Commun. 2021 Nov 1;12(1):6271. doi: 10.1038/s41467-021-26571-7. Nat Commun. 2021. PMID: 34725346 Free PMC article.
-
Molecular mechanism of methyl-dependent and spatial-specific DNA recognition of c-Jun homodimer.J Mol Model. 2021 Jul 15;27(8):227. doi: 10.1007/s00894-021-04840-y. J Mol Model. 2021. PMID: 34264385
-
Energetic coupling along an allosteric communication channel drives the binding of Jun-Fos heterodimeric transcription factor to DNA.FEBS J. 2011 Jun;278(12):2090-104. doi: 10.1111/j.1742-4658.2011.08124.x. Epub 2011 May 18. FEBS J. 2011. PMID: 21496208 Free PMC article.
References
-
- Chinenov Y, Kerppola TK. Close encounters of many kinds: Fos-Jun interactions that mediate transcription regulatory specificity. Oncogene. 2001;20:2438–2452. - PubMed
-
- Angel P, Karin M. The role of Jun, Fos and the AP-1 complex in cell-proliferation and transformation. Biochim Biophys Acta. 1991;1072:129–157. - PubMed
-
- Baxevanis AD, Vinson CR. Interactions of coiled coils in transcription factors: where is the specificity? Curr Opin Genet Dev. 1993;3:278–285. - PubMed
-
- Raivich G, Behrens A. Role of the AP-1 transcription factor c-Jun in developing, adult and injured brain. Prog Neurobiol. 2006;78:347–363. - PubMed
-
- Milde-Langosch K. The Fos family of transcription factors and their role in tumourigenesis. Eur J Cancer. 2005;41:2449–2461. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
