

A dynamin is required for the biogenesis of secretory organelles in Toxoplasma gondii
- PMID: 19217293
- PMCID: PMC3941470
- DOI: 10.1016/j.cub.2009.01.039
A dynamin is required for the biogenesis of secretory organelles in Toxoplasma gondii
Abstract
Background: Apicomplexans contain only a core set of factors involved in vesicular traffic. Yet these obligate intracellular parasites evolved a set of unique secretory organelles (micronemes, rhoptries, and dense granules) that are required for invasion and modulation of the host cell. Apicomplexa replicate by budding from or within a single mother cell, and secretory organelles are synthesized de novo at the final stage of division. To date, the molecular basis for their biogenesis is unknown.
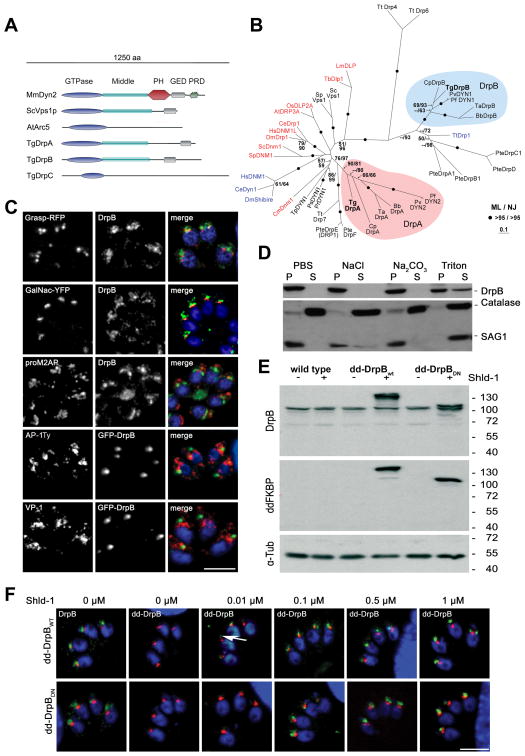
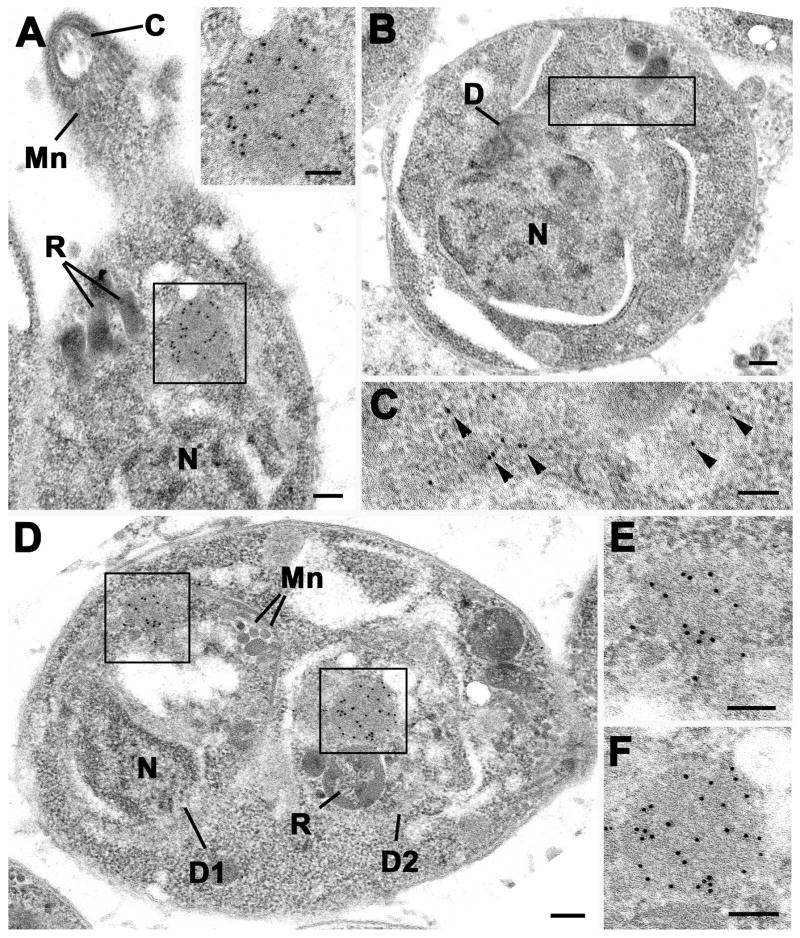
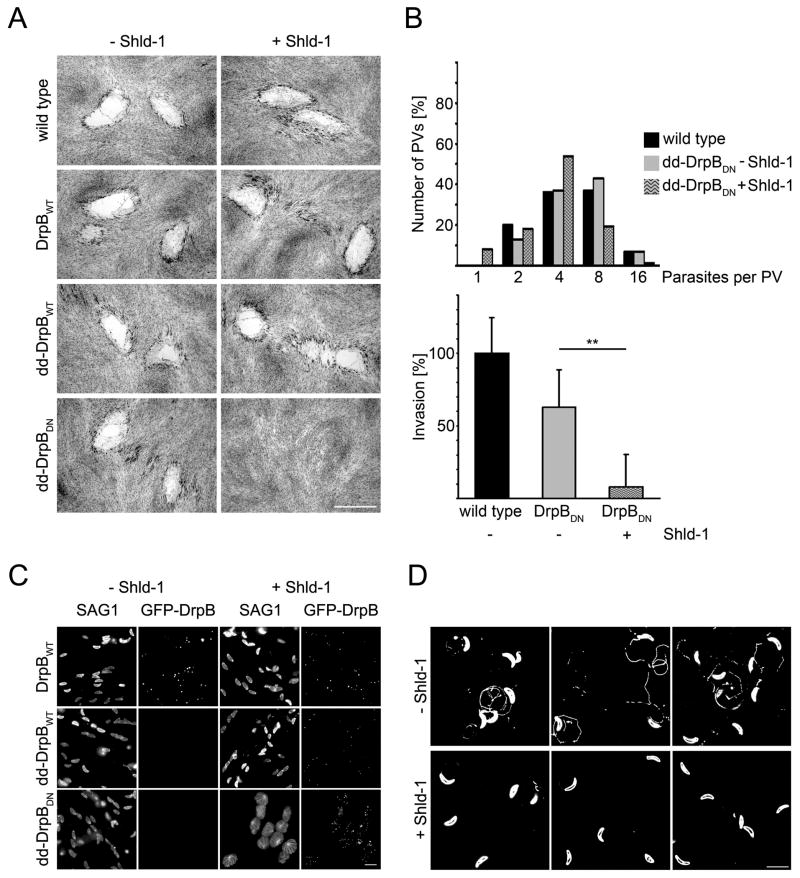
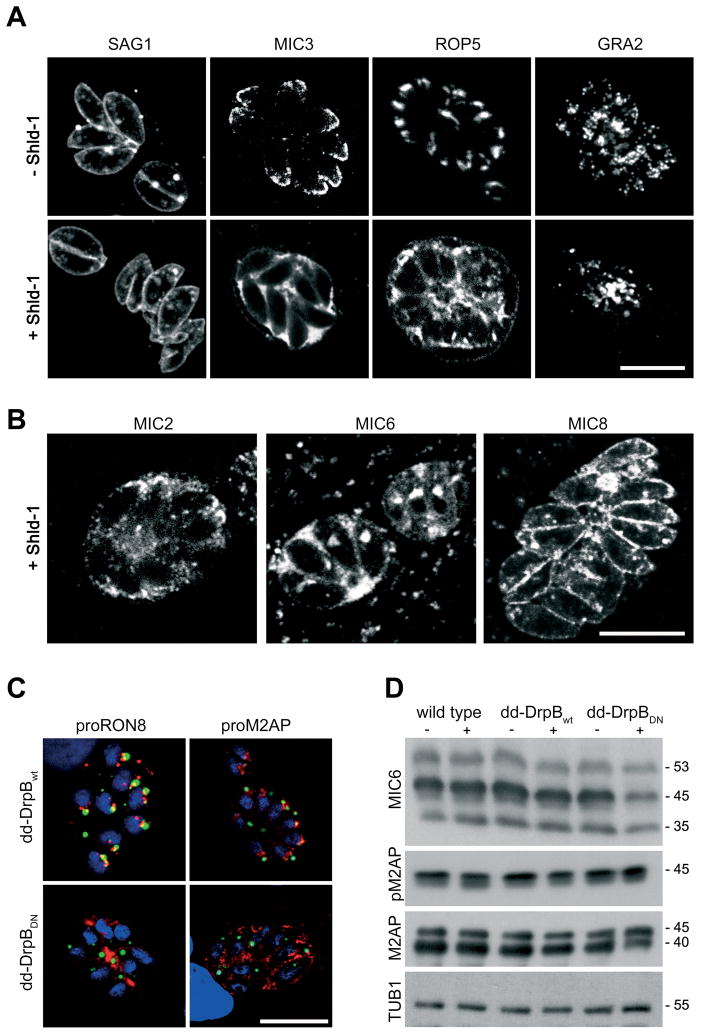
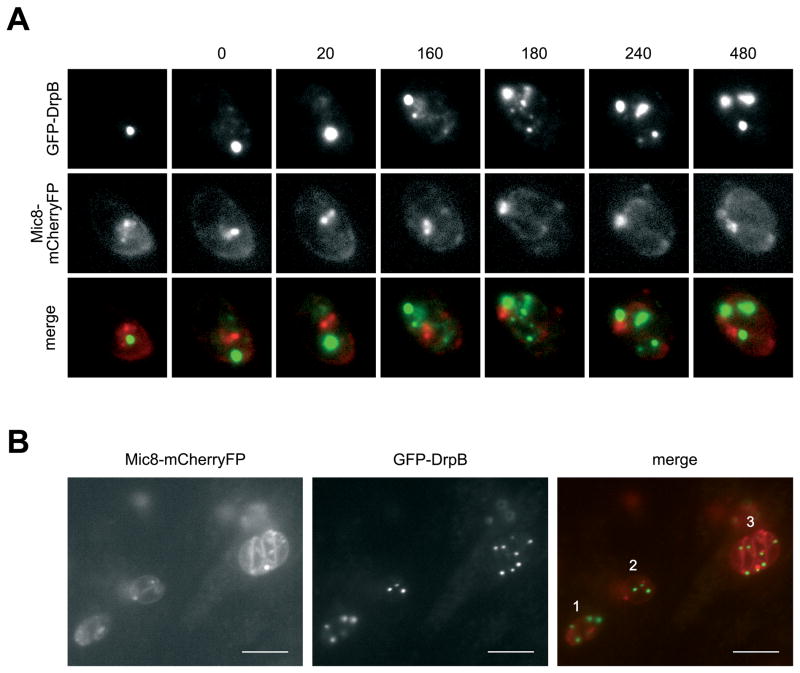
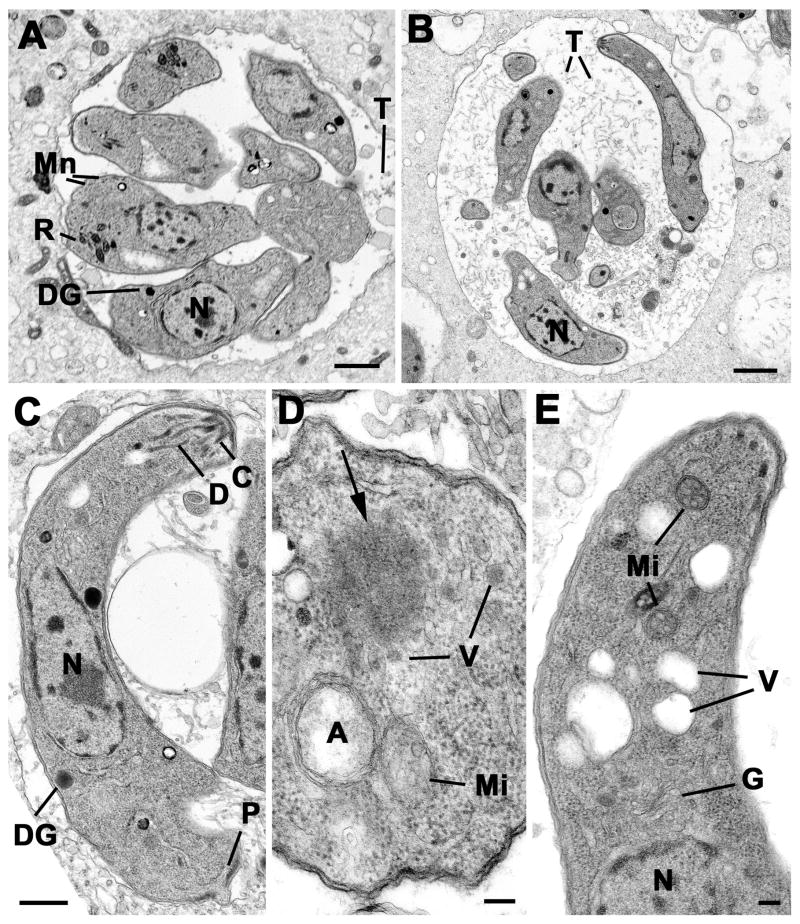
Results: We demonstrate that the apicomplexan dynamin-related protein B (DrpB) belongs to an alveolate specific family of dynamins that is expanded in ciliates. DrpB accumulates in a cytoplasmic region close to the Golgi that breaks up during replication and reforms after assembly of the daughter cells. Conditional ablation of DrpB function results in mature daughter parasites that are devoid of micronemes and rhoptries. In the absence of these organelles, invasion-related secretory proteins are mistargeted to the constitutive secretory pathway. Mutant parasites are able to replicate but are unable to escape from or invade into host cells.
Conclusions: DrpB is the essential mechanoenzyme for the biogenesis of secretory organelles in Apicomplexa. We suggest that DrpB is required during replication to generate vesicles for the regulated secretory pathway that form the unique secretory organelles. Our study supports a role of an alveolate-specific dynamin that was required for the evolution of novel, secretory organelles. In the case of Apicomplexa, these organelles further evolved to enable a parasitic lifestyle.
Figures
Comment in
-
Organelle division: dynamin-related proteins in apicomplexans.Curr Biol. 2009 Apr 28;19(8):R334-6. doi: 10.1016/j.cub.2009.02.044. Curr Biol. 2009. PMID: 19409284
References
-
- Carruthers V, Boothroyd JC. Pulling together: an integrated model of Toxoplasma cell invasion. Curr Opin Microbiol. 2007;10:83–89. - PubMed
-
- Sheiner L, Soldati-Favre D. Protein trafficking inside Toxoplasma gondii. Traffic. 2008;9:636–646. - PubMed
-
- Ngo HM, Yang M, Paprotka K, Pypaert M, Hoppe H, Joiner KA. AP-1 in Toxoplasma gondii mediates biogenesis of the rhoptry secretory organelle from a post-Golgi compartment. J Biol Chem. 2003;278:5343–5352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
