

Review
doi: 10.1016/j.cbpa.2008.12.018.
Epub 2009 Feb 11.
Revisiting the modularity of modular polyketide synthases
Affiliations
- PMID: 19217343
- PMCID: PMC2737389
- DOI: 10.1016/j.cbpa.2008.12.018
Item in Clipboard
Review
Revisiting the modularity of modular polyketide synthases
Curr Opin Chem Biol.
2009 Apr.
Abstract
Modularity is a highly sought after feature in engineering design. A modular catalyst is a multi-component system whose parts can be predictably interchanged for functional flexibility and variety. Nearly two decades after the discovery of the first modular polyketide synthase (PKS), we critically assess PKS modularity in the face of a growing body of atomic structural and in vitro biochemical investigations. Both the architectural modularity and the functional modularity of this family of enzymatic assembly lines are reviewed, and the fundamental challenges that lie ahead for the rational exploitation of their full biosynthetic potential are discussed.
Figures
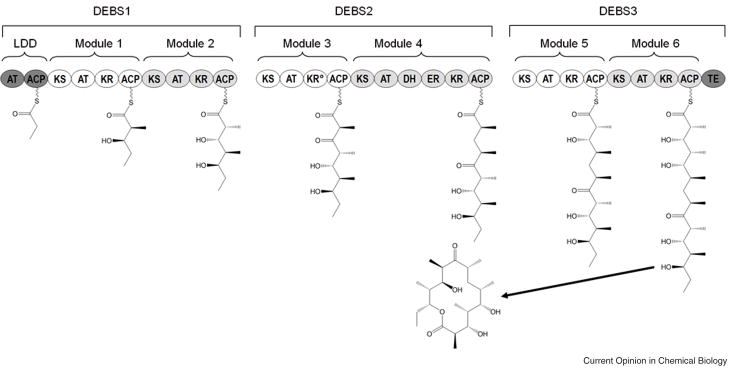
Chain elongation occurs minimally through the combined action of the ketosynthase (KS), acyl transferase (AT), and acyl carrier protein (ACP) domains. The final oxidation state of the ®-carbon is controlled by the specific combination of ketoreductase (KR), dehydratase (DH) and enoylreductase (ER) domains present in a given module. Once processed, the polyketide chain is either passed to the KS domain of the downstream module or cyclized and released by the thioesterase (TE) domain at the C-terminus of the polyketide synthase. The loading didomain (LDD) is responsible for the selection and subsequent loading of the appropriate priming unit. KR°, inactive ketoreductase domain.

Orange, N-terminal coiled coil linker that facilitates docking between modules 4 and 5; blue, ketosynthase (KS) domain; green, acyltransferase (AT) domain; yellow, KS-AT linker; red, AT-KR linker; cyan, ketoreductase (KR) domain; magenta, acyl carrier protein (ACP) domain. (Only a single ACP domain is shown for clarity.) The sequences (colored by domain/linker of origin) of junctions at which the KS, AT, KR and ACP domains can be deconstructed into stand-alone proteins, or alternatively recombined, are indicated with black arrows. The latter half of the AT-KR linker (red, residues 902–911) and the KR-ACP linker (black, residues 1360–1377) are not shown, as there is no reliable data based on which they can be modeled. The relative orientation of the ACP and KS domains is predicted by in silico docking analysis, whereas the relative orientation of the KR domains and the remainder of the module is based on the X-ray structure of homologous porcine fatty acid synthase [17].
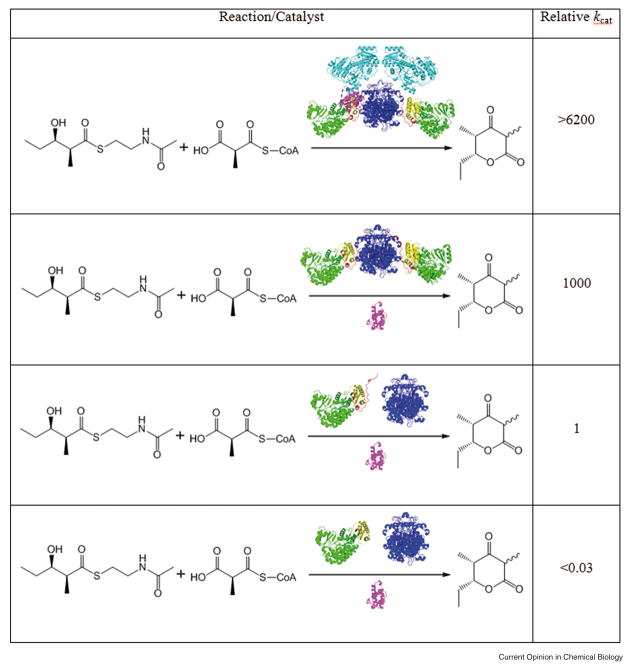
Domains and linker regions are colored as in Figure 2. The relative turnover number of the intact module (first entry) is compared with that of a derivative in which: (i) a stand-alone ACP is used (second entry); (ii) the KS and AT are discrete proteins (third entry); or (iii) an analog of (ii) lacking the post-AT linker (fourth entry). In the third and fourth entries, only the KS is shown as a dimer.

A 1000-fold variation in specificity is observed, with both kcat and kcat/KM values decreasing as the natural ACP domain (ACP3) is replaced by the ACP domains from modules 1, 2, 5 and 6 of DEBS. Data taken from [32••].


References
-
- Yoon TP, Jacobsen EN. Privileged chiral catalysts. Science. 2003;299:1691–1693. - PubMed
-
- Burk MJ. Modular Phospholane Ligands in Asymmetric Catalysis. Acc Chem Res. 2000;33:363–372. - PubMed
-
- Menzella HG, Reid R, Carney JR, Chandran SS, Reisinger SJ, Patel KG, Hopwood DA, Santi DV. Combinatorial polyketide biosynthesis by de novo design and rearrangement of modular polyketide synthase genes. Nat Biotechnol. 2005;23:1171–1176. - PubMed
-
- Kao CM, McPherson M, McDaniel R, Fu H, Cane DE, Khosla C. Alcohol stereochemistry in polyketide backbones is controlled by the beta-ketoreductase domains of modular polyketide synthases. J Am Chem Soc. 1998;120:2478–2479.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
