

Molecular architecture of the human pre-mRNA 3' processing complex
- PMID: 19217410
- PMCID: PMC2946185
- DOI: 10.1016/j.molcel.2008.12.028
Molecular architecture of the human pre-mRNA 3' processing complex
Abstract
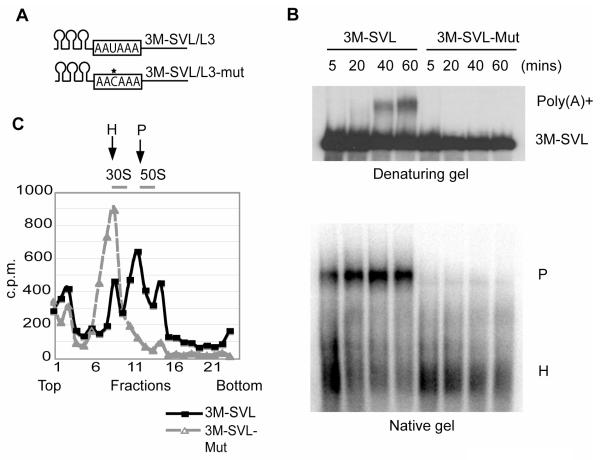
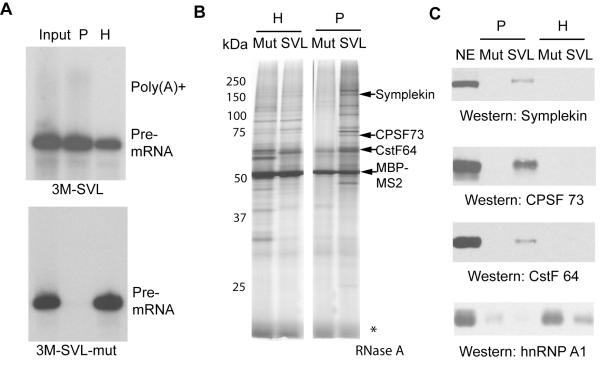
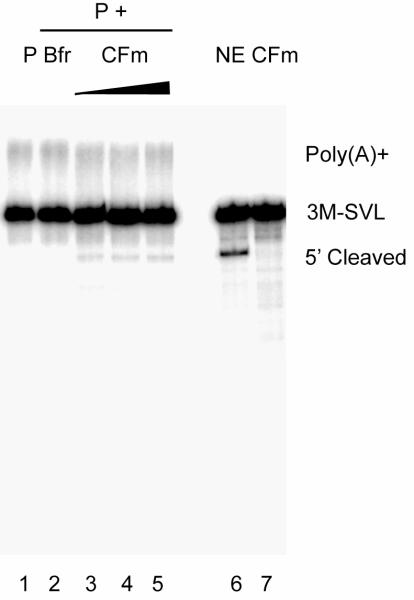
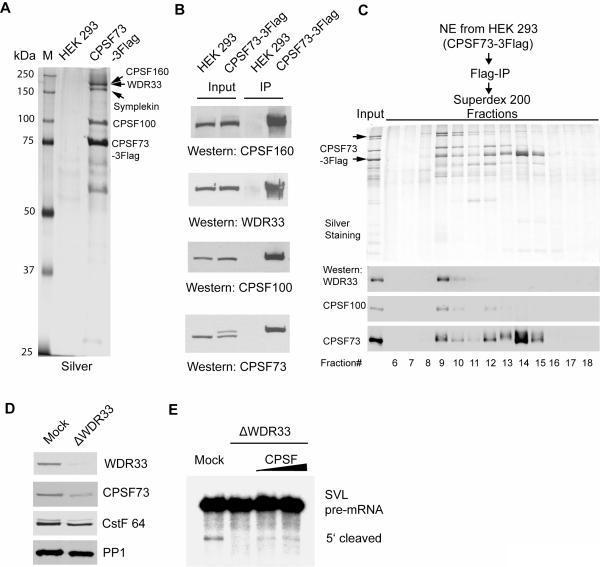
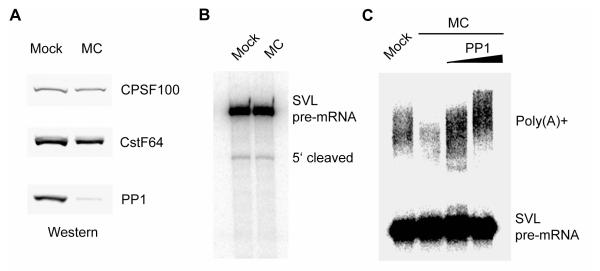
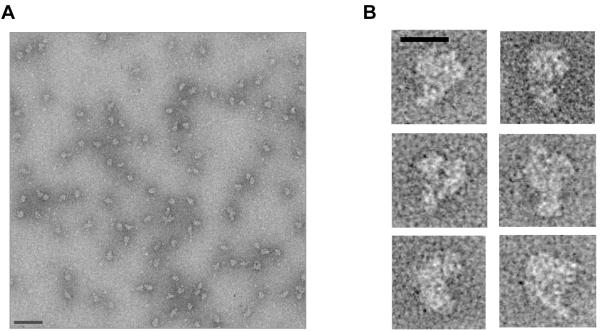
Pre-mRNA 3' end formation is an essential step in eukaryotic gene expression. Over half of human genes produce alternatively polyadenylated mRNAs, suggesting that regulated polyadenylation is an important mechanism for posttranscriptional gene control. Although a number of mammalian mRNA 3' processing factors have been identified, the full protein composition of the 3' processing machinery has not been determined, and its structure is unknown. Here we report the purification and subsequent proteomic and structural characterization of human mRNA 3' processing complexes. Remarkably, the purified 3' processing complex contains approximately 85 proteins, including known and new core 3' processing factors and over 50 proteins that may mediate crosstalk with other processes. Electron microscopic analyses show that the core 3' processing complex has a distinct "kidney" shape and is approximately 250 A in length. Together, our data has revealed the complexity and molecular architecture of the pre-mRNA 3' processing complex.
Figures
References
-
- Bai Y, Auperin TC, Chou CY, Chang GG, Manley JL, Tong L. Crystal structure of murine CstF-77: dimeric association and implications for polyadenylation of mRNA precursors. Mol Cell. 2007;25:863–875. - PubMed
-
- Baillat D, Hakimi MA, Naar AM, Shilatifard A, Cooch N, Shiekhattar R. Integrator, a multiprotein mediator of small nuclear RNA processing, associates with the C-terminal repeat of RNA polymerase II. Cell. 2005;123:265–276. - PubMed
-
- Bard J, Zhelkovsky AM, Helmling S, Earnest TN, Moore CL, Bohm A. Structure of yeast poly(A) polymerase alone and in complex with 3′-dATP. Science. 2000;289:1346–1349. - PubMed
-
- Bentley DL. Rules of engagement: co-transcriptional recruitment of pre-mRNA processing factors. Curr Opin Cell Biol. 2005;17:251–256. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
