

Interactions with the Abelson tyrosine kinase reveal compartmentalization of eyes absent function between nucleus and cytoplasm
- PMID: 19217428
- PMCID: PMC2670569
- DOI: 10.1016/j.devcel.2008.12.005
Interactions with the Abelson tyrosine kinase reveal compartmentalization of eyes absent function between nucleus and cytoplasm
Abstract
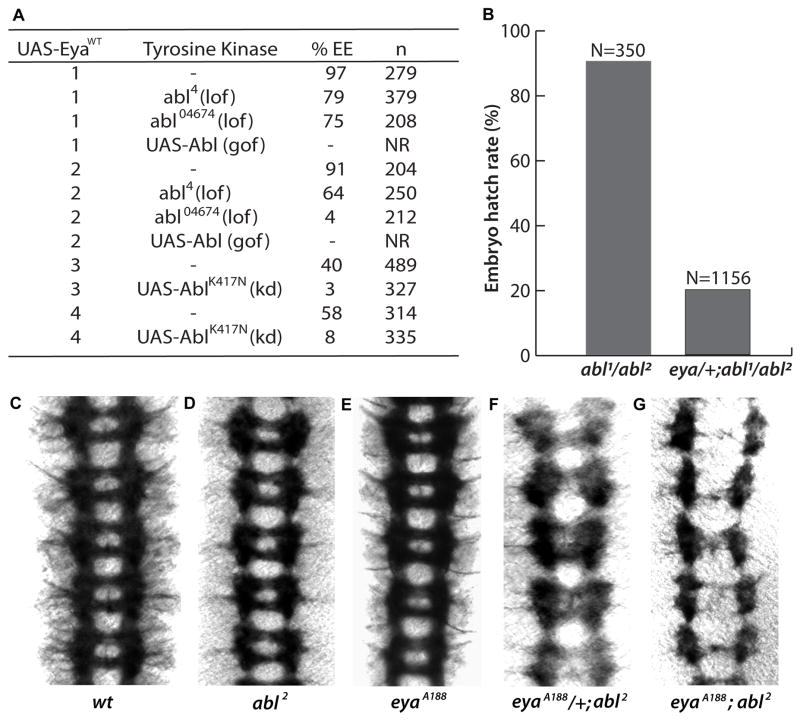
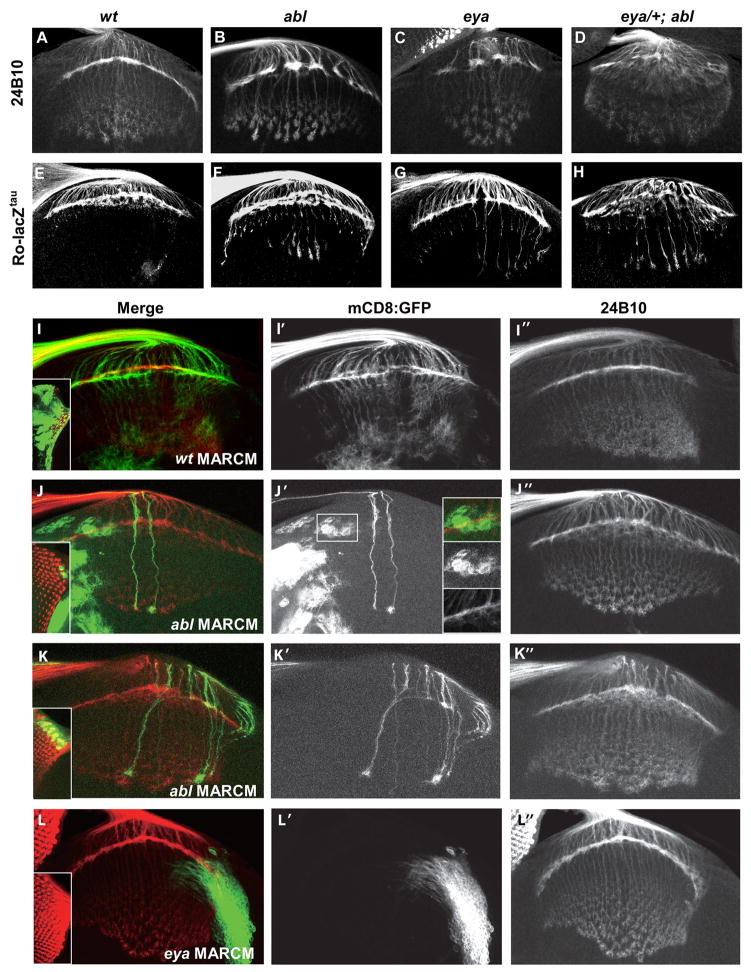
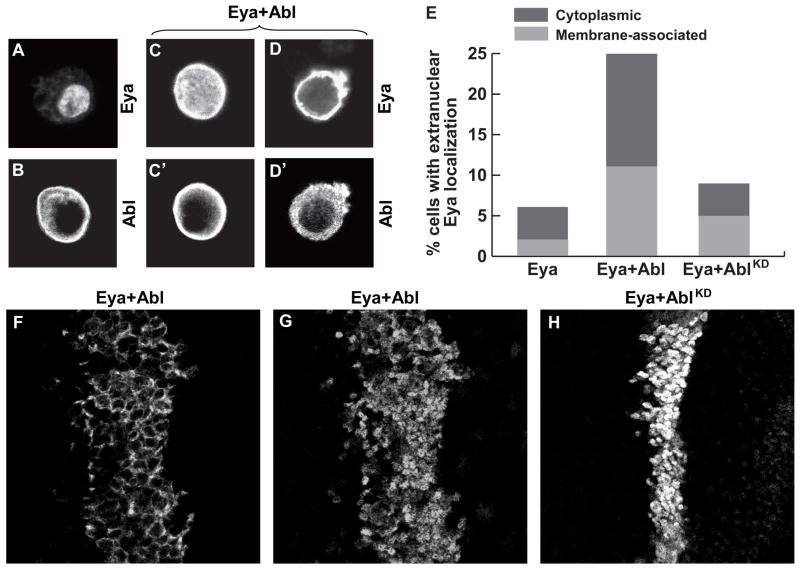
Eyes absent (Eya), named for its role in Drosophila eye development but broadly conserved in metazoa, possesses dual functions as a transcriptional coactivator and protein tyrosine phosphatase. Although Eya's transcriptional activity has been extensively characterized, the physiological requirements for its phosphatase activity remain obscure. In this study, we provide insight into Eya's participation in phosphotyrosine-mediated signaling networks by demonstrating cooperative interactions between Eya and the Abelson (Abl) tyrosine kinase during development of the Drosophila larval visual system. Mechanistically, Abl-mediated phosphorylation recruits Eya to the cytoplasm, where in vivo studies reveal a requirement for its phosphatase function. Thus, we propose a model in which, in addition to its role as a transcription factor, Eya functions as a cytoplasmic protein tyrosine phosphatase.
Figures



References
-
- Abdelhak S, Kalatzis V, et al. A human homologue of the Drosophila eyes absent gene underlies branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nat Genet. 1997;15(2):157–64. - PubMed
-
- Bennett RL, Hoffmann FM. Increased levels of the Drosophila Abelson tyrosine kinase in nerves and muscles: subcellular localization and mutant phenotypes imply a role in cell-cell interactions. Development. 1992;116(4):953–66. - PubMed
-
- Bonini NM, Bui QT, et al. The Drosophila eyes absent gene directs ectopic eye formation in a pathway conserved between flies and vertebrates. Development. 1997;124(23):4819–26. - PubMed
-
- Bonini NM, Leiserson WM, et al. The eyes absent gene: genetic control of cell survival and differentiation in the developing Drosophila eye. Cell. 1993;72(3):379–95. - PubMed
-
- Bonini NM, Leiserson WM, et al. Multiple roles of the eyes absent gene in Drosophila. Dev Biol. 1998;196(1):42–57. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
