

Displacement of formins from growing barbed ends by bud14 is critical for actin cable architecture and function
- PMID: 19217430
- PMCID: PMC2667650
- DOI: 10.1016/j.devcel.2008.12.001
Displacement of formins from growing barbed ends by bud14 is critical for actin cable architecture and function
Abstract
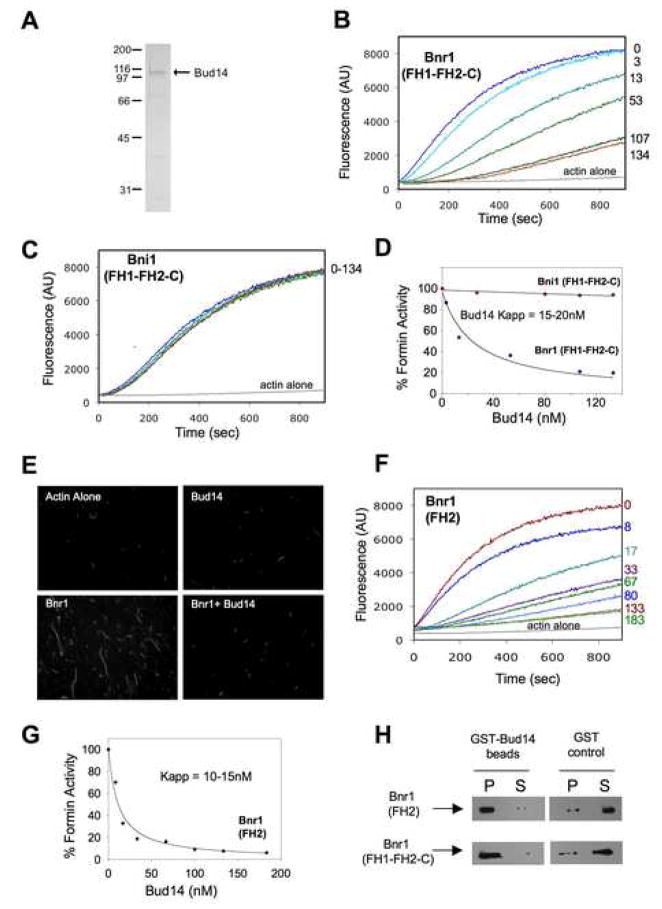
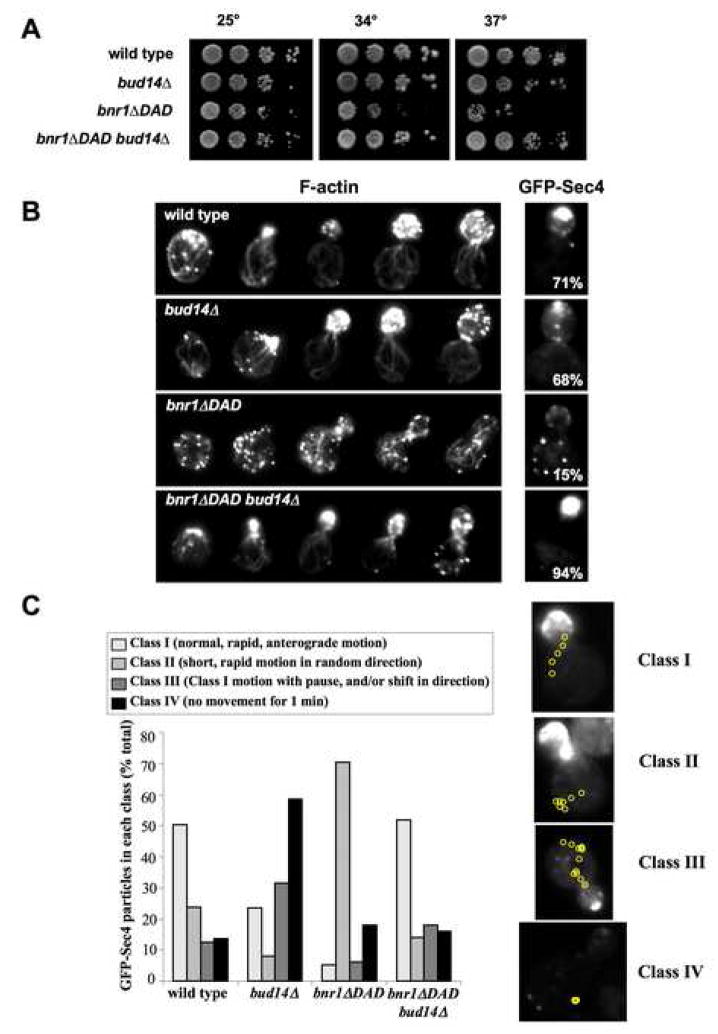
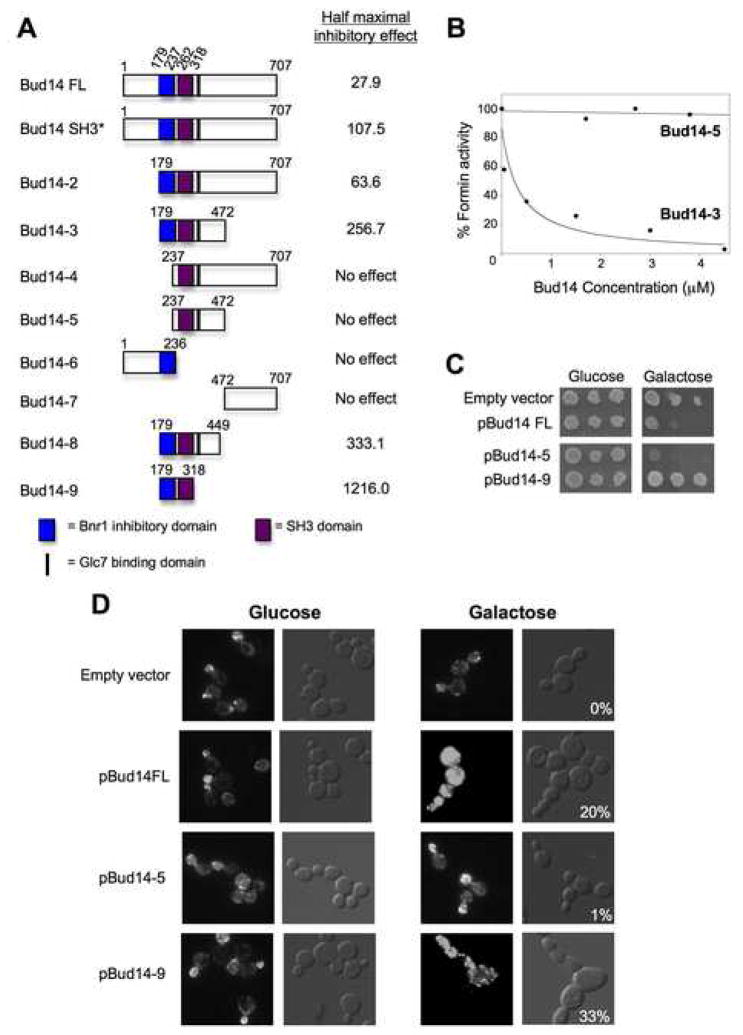
Normal cellular development and function require tight spatiotemporal control of actin assembly. Formins are potent actin assembly factors that protect the growing ends of actin filaments from capping proteins. However, it is unresolved how the duration of formin-mediated actin assembly events is controlled, whether formins are actively displaced from growing ends, and how filament length is regulated in vivo. Here, we identify Bud14 as a high-affinity inhibitor of the yeast formin Bnr1 that rapidly displaces the Bnr1 FH2 domain from growing barbed ends. Consistent with these activities, bud14Delta cells display fewer actin cables, which are aberrantly long, bent, and latrunculinA resistant, leading to defects in secretory vesicle movement. Moreover, bud14Delta suppressed mutations that cause abnormally numerous and shortened cables, restoring wild-type actin architecture. From these results, we propose that formin displacement factors regulate filament length and are required in vivo to maintain proper actin network architecture and function.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
