

14-3-3:Shc scaffolds integrate phosphoserine and phosphotyrosine signaling to regulate phosphatidylinositol 3-kinase activation and cell survival
- PMID: 19218246
- PMCID: PMC2673277
- DOI: 10.1074/jbc.M807637200
14-3-3:Shc scaffolds integrate phosphoserine and phosphotyrosine signaling to regulate phosphatidylinositol 3-kinase activation and cell survival
Abstract
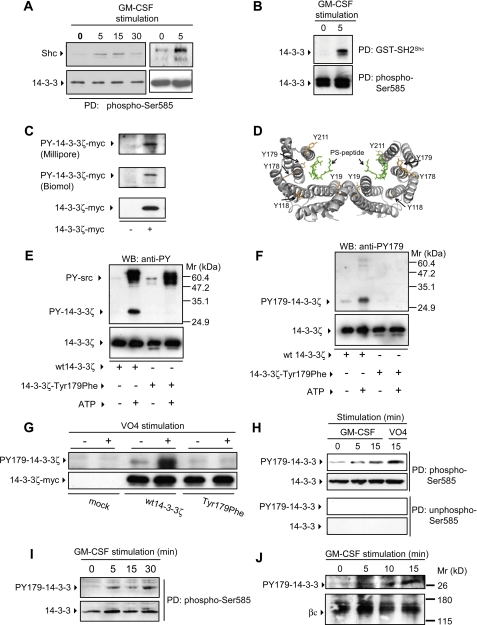
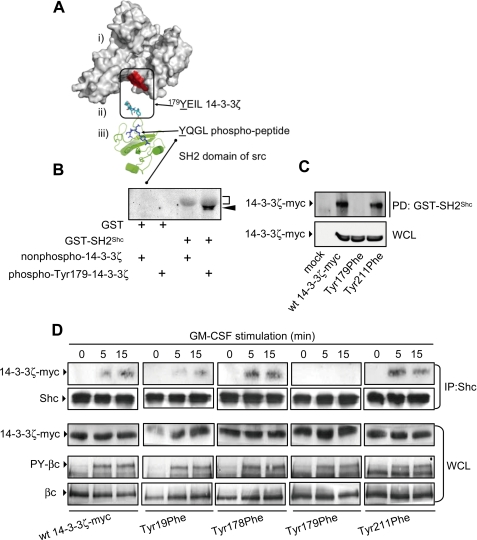
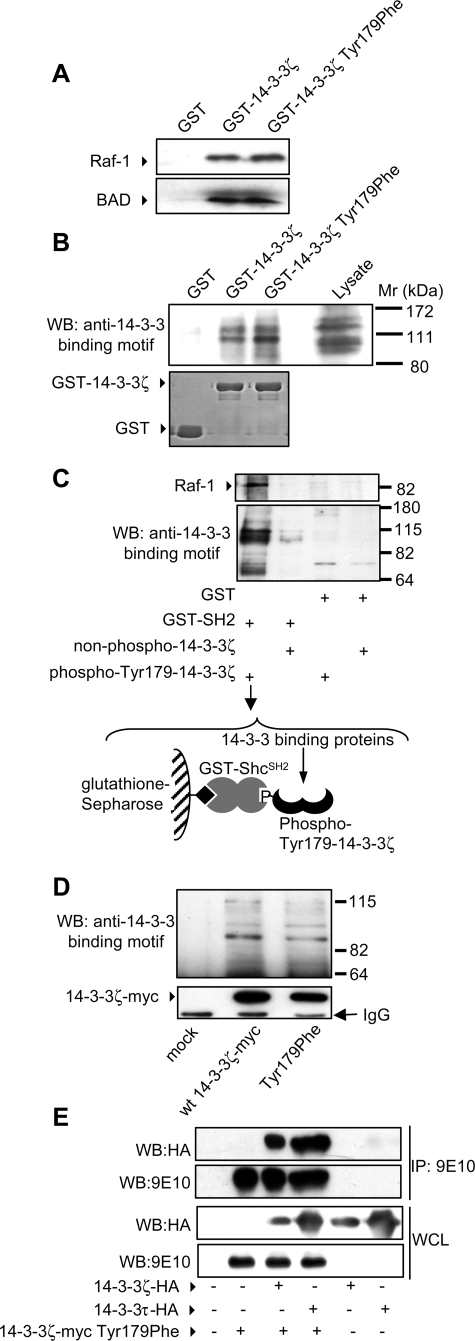
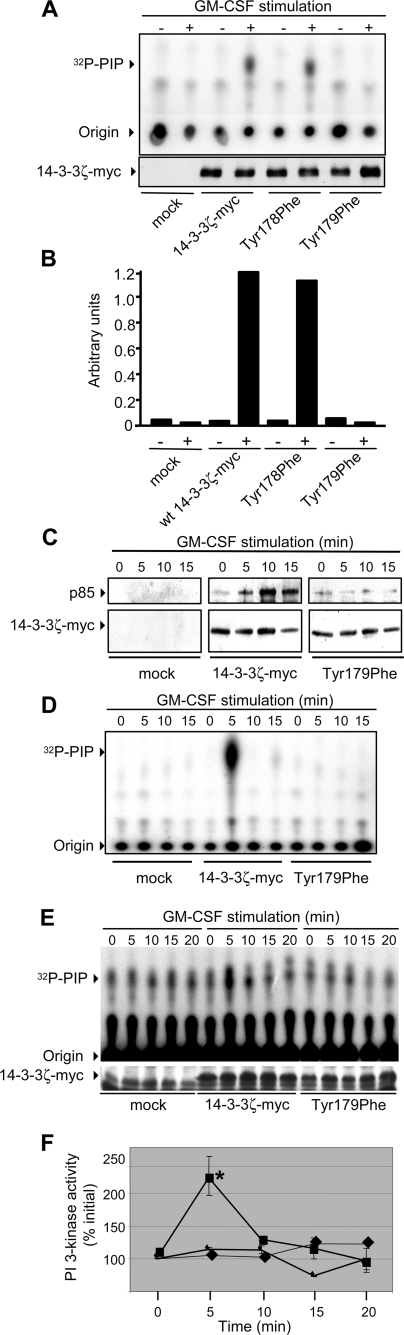
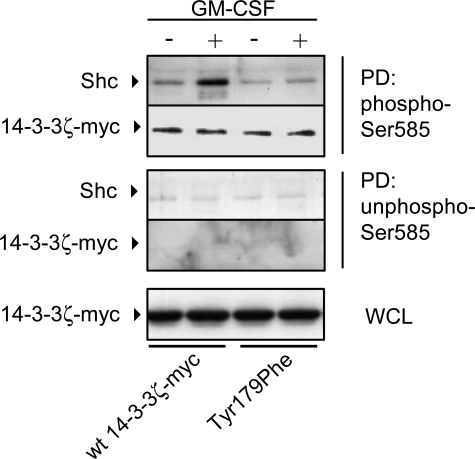
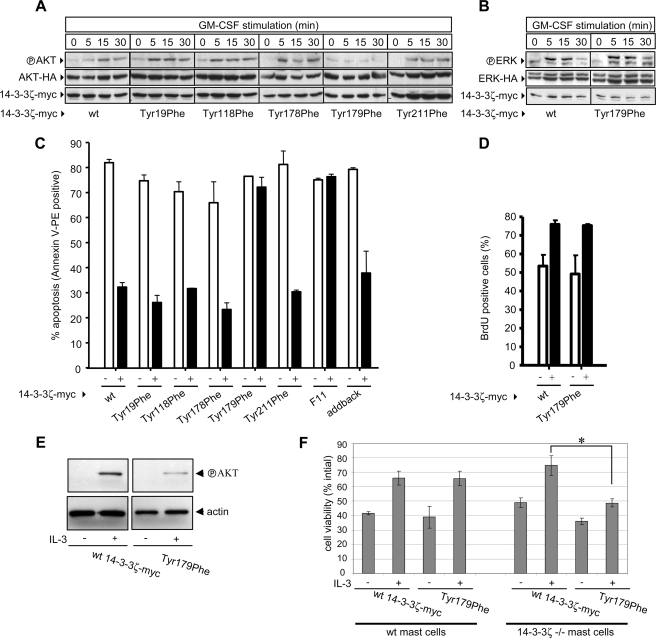
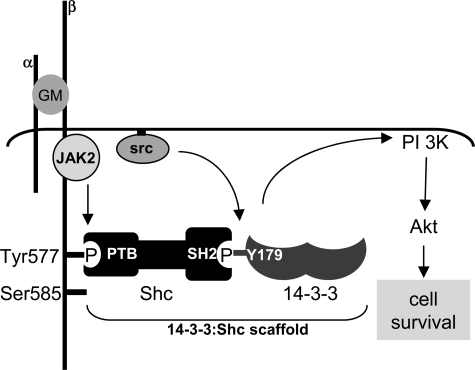
Integrated cascades of protein tyrosine and serine/threonine phosphorylation play essential roles in transducing signals in response to growth factors and cytokines. How adaptor or scaffold proteins assemble signaling complexes through both phosphotyrosine and phosphoserine/threonine residues to regulate specific signaling pathways and biological responses is unclear. We show in multiple cell types that endogenous 14-3-3zeta is phosphorylated on Tyr(179) in response to granulocyte macrophage colony-stimulating factor. Importantly, 14-3-3zeta can function as an intermolecular bridge that couples to phosphoserine residues and also directly binds the SH2 domain of Shc via Tyr(179). The assembly of these 14-3-3:Shc scaffolds is specifically required for the recruitment of a phosphatidylinositol 3-kinase signaling complex and the regulation of CTL-EN cell survival in response to cytokine. The biological significance of these findings was further demonstrated using primary bone marrow-derived mast cells from 14-3-3zeta(-/-) mice. We show that cytokine was able to promote Akt phosphorylation and viability of primary mast cells derived from 14-3-3zeta(-/-) mice when reconstituted with wild type 14-3-3zeta, but the Akt phosphorylation and survival response was reduced in cells reconstituted with the Y179F mutant. Together, these results show that 14-3-3:Shc scaffolds can act as multivalent signaling nodes for the integration of both phosphoserine/threonine and phosphotyrosine pathways to regulate specific cellular responses.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
