

Time-resolved photolabeling of the nicotinic acetylcholine receptor by [3H]azietomidate, an open-state inhibitor
- PMID: 19218367
- PMCID: PMC2672814
- DOI: 10.1124/mol.108.054353
Time-resolved photolabeling of the nicotinic acetylcholine receptor by [3H]azietomidate, an open-state inhibitor
Abstract
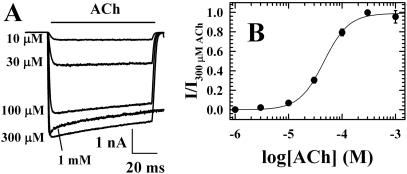
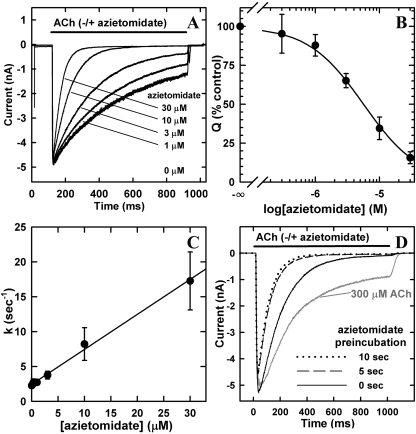
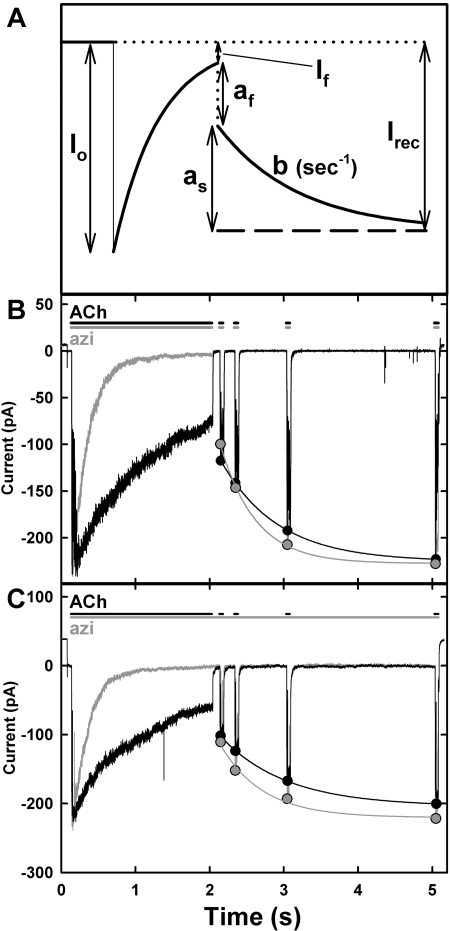
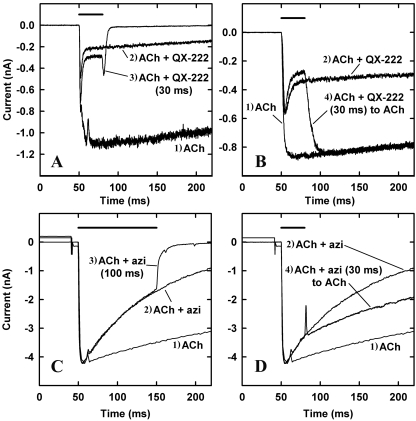
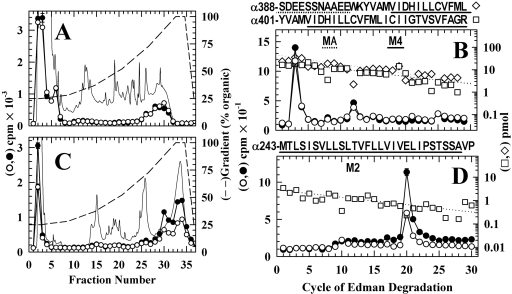
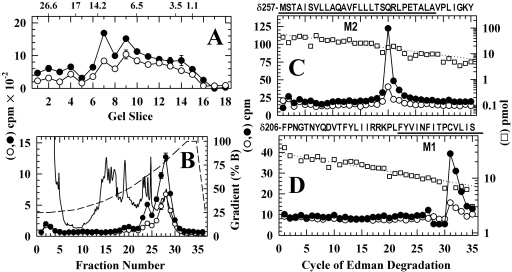
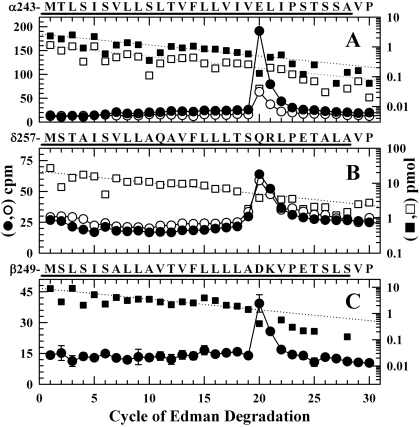
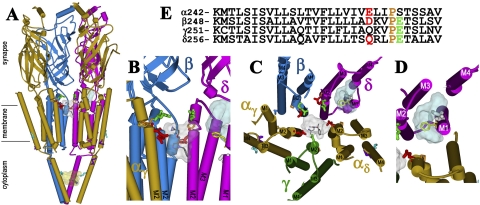
Azietomidate is a photoreactive analog of the general anesthetic etomidate that acts as a nicotinic acetylcholine receptor (nAChR) noncompetitive antagonist. We used rapid perfusion electrophysiological techniques to characterize the state dependence and kinetics of azietomidate inhibition of Torpedo californica nAChRs and time-resolved photolabeling to identify the nAChR binding sites occupied after exposure to [(3)H]azietomidate and agonist for 50 ms (open state) or at equilibrium (desensitized state). Azietomidate acted primarily as an open channel inhibitor characterized by a bimolecular association rate constant of k(+) = 5 x 10(5) M(-1) s(-1) and a dissociation rate constant of <3s(-1). Azietomidate at 10 microM, when perfused with acetylcholine (ACh), inhibited the ACh response by approximately 50% after 50 ms; when preincubated for 10 s, it decreased the peak initial response by approximately 15%. Comparison of the kinetics of recovery of ACh responses after exposure to ACh and azietomidate or to ACh alone indicated that at subsecond times, azietomidate inhibited nAChRs without enhancing the kinetics of agonist-induced desensitization. In nAChRs frozen after 50-ms exposure to agonist and [(3)H]azietomidate, amino acids were photolabeled in the ion channel [position M2-20 (alphaGlu-262, betaAsp-268, deltaGln-276)], in deltaM1 (deltaCys-236), and in alphaMA/alphaM4 (alphaGlu-390, alphaCys-412) that were also photolabeled in nAChRs in the equilibrium desensitized state at approximately half the efficiency. These results identify azietomidate binding sites at the extracellular end of the ion channel, in the delta subunit helix bundle, and in the nAChR cytoplasmic domain that seem similar in structure and accessibility in the open and desensitized states of the nAChR.
Figures
References
-
- Addona GH, Kloczewiak MA, and Miller KW (1999) Time-resolved photolabeling of membrane proteins: application to the nicotinic acetylcholine receptor. Anal Biochem 267 135-140. - PubMed
-
- Arevalo E, Chiara DC, Forman SA, Cohen JB, and Miller KW (2005) Gating-enhanced accessibility of hydrophobic sites within the transmembrane region of the nicotinic acetylcholine receptor's delta-subunit - a time-resolved photolabeling study. J Biol Chem 280 13631-13640. - PubMed
-
- Arias HR, Kem WR, Trudell JR, and Blanton MP (2003) Unique general anesthetic binding sites within distinct conformational states of the nicotinic acetylcholine receptor. Int Rev Neurobiol 54 1-50. - PubMed
-
- Borghese CM, Ali DN, Bleck V, and Harris RA (2002) Acetylcholine and alcohol sensitivity of neuronal nicotinic acetylcholine receptors: mutations in transmembrane domains. Alcohol Clin Exp Res 26 1764-1772. - PubMed
-
- Chiara DC, Trinidad JC, Wang D, Ziebell MR, Sullivan D, and Cohen JB (2003) Identification of amino acids in the nicotinic acetylcholine receptor agonist binding site and ion channel photolabeled by 4-[(3-trifluoromethyl)-3H-diazirin-3-yl]benzoyllcholine, a novel photoaffinity antagonist. Biochemistry 42 271-283. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
