

Nuclear activity of ROXY1, a glutaredoxin interacting with TGA factors, is required for petal development in Arabidopsis thaliana
- PMID: 19218396
- PMCID: PMC2660636
- DOI: 10.1105/tpc.108.064477
Nuclear activity of ROXY1, a glutaredoxin interacting with TGA factors, is required for petal development in Arabidopsis thaliana
Erratum in
- Plant Cell. 2014 Mar;26(3):1378
-
CORRECTION.Plant Cell. 2018 Jan;30(1):249-252. doi: 10.1105/tpc.17.00893. Epub 2018 Jan 12. Plant Cell. 2018. PMID: 29330201 Free PMC article. No abstract available.
Abstract
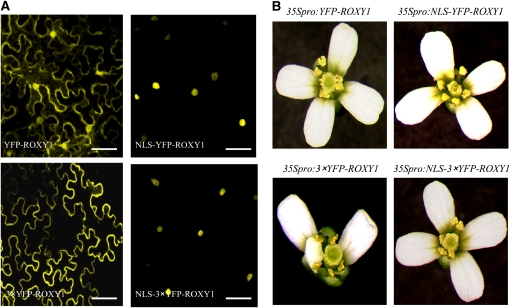
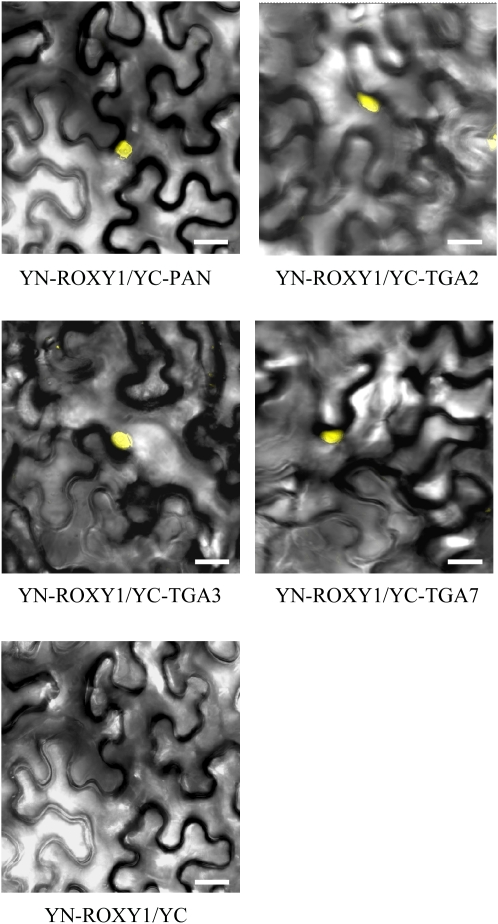
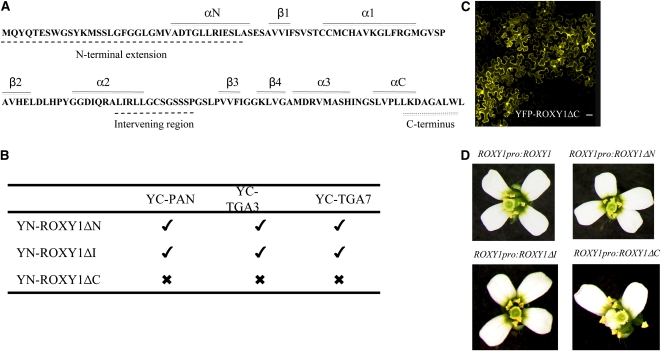
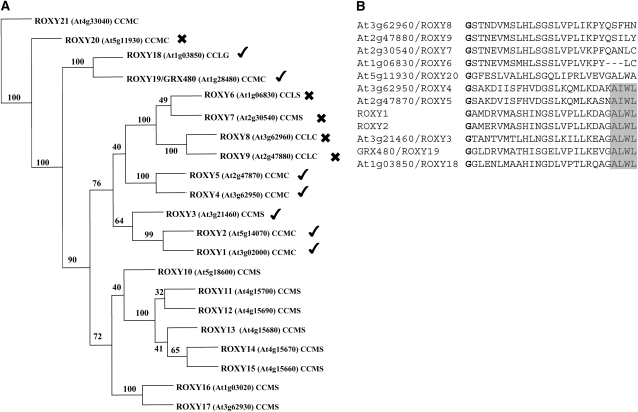
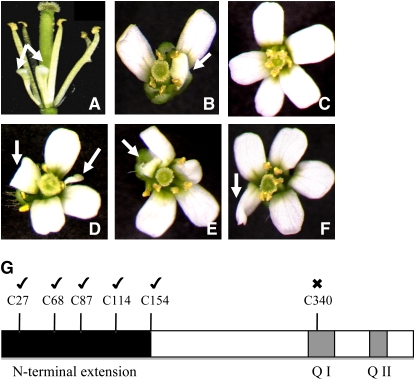
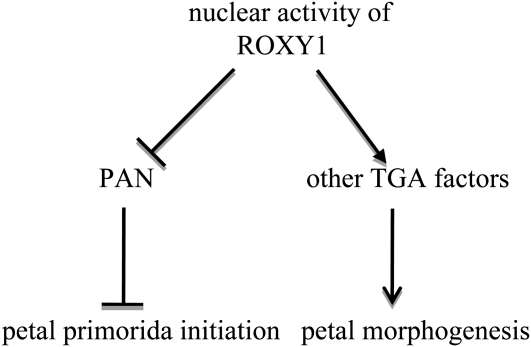
Glutaredoxins (GRXs) have thus far been associated mainly with redox-regulated processes participating in stress responses. However, ROXY1, encoding a GRX, has recently been shown to regulate petal primorida initiation and further petal morphogenesis in Arabidopsis thaliana. ROXY1 belongs to a land plant-specific class of GRXs that has a CC-type active site motif, which deviates from ubiquitously occurring CPYC and CGFS GRXs. Expression studies of yellow fluorescent protein-ROXY1 fusion genes driven by the cauliflower mosaic virus 35S promoter reveal a nucleocytoplasmic distribution of ROXY1. We demonstrate that nuclear localization of ROXY1 is indispensable and thus crucial for its activity in flower development. Yeast two-hybrid screens identified TGA transcription factors as interacting proteins, which was confirmed by bimolecular fluorescence complementation experiments showing their nuclear interaction in planta. Overlapping expression patterns of ROXY1 and TGA genes during flower development demonstrate that ROXY1/TGA protein interactions can occur in vivo and support their biological relevance in petal development. Deletion analysis of ROXY1 demonstrates the importance of the C terminus for its functionality and for mediating ROXY1/TGA protein interactions. Phenotypic analysis of the roxy1-2 pan double mutant and an engineered chimeric repressor mutant from PERIANTHIA (PAN), a floral TGA gene, supports a dual role of ROXY1 in petal development. Together, our results show that the ROXY1 protein functions in the nucleus, likely by modifying PAN posttranslationally and thereby regulating its activity in petal primordia initiation. Additionally, ROXY1 affects later petal morphogenesis, probably by modulating other TGA factors that might act redundantly during differentiation of second whorl organs.
Figures


References
-
- Bandyopadhyay, S., Gama, F., Molina-Navarro, M.M., Gualberto, J.M., Claxton, R., Naik, S.G., Huynh, B.H., Herrero, E., Jacquot, J.P., Johnson, M.K., and Rouhier, N. (2008). Chloroplast monothiol glutaredoxins as scaffold proteins for the assembly and delivery of [2Fe-2S] clusters. EMBO J. 27 1122–1133. - PMC - PubMed
-
- Buchanan, B.B., and Balmer, Y. (2005). Redox regulation: A broadening horizon. Annu. Rev. Plant Biol. 56 187–220. - PubMed
-
- Cheng, N.H., Liu, J.Z., Brock, A., Nelson, R.S., and Hirschi, K.D. (2006). AtGRXcp, an Arabidopsis chloroplastic glutaredoxin, is critical for protection against protein oxidative damage. J. Biol. Chem. 281 26280–26288. - PubMed
-
- Clough, S.J., and Bent, A.F. (1998). Floral dip: A simplified method for Agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J. 16 735–743. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
