

Mechanistic insights into active site-associated polyubiquitination by the ubiquitin-conjugating enzyme Ube2g2
- PMID: 19223579
- PMCID: PMC2644258
- DOI: 10.1073/pnas.0808564106
Mechanistic insights into active site-associated polyubiquitination by the ubiquitin-conjugating enzyme Ube2g2
Abstract
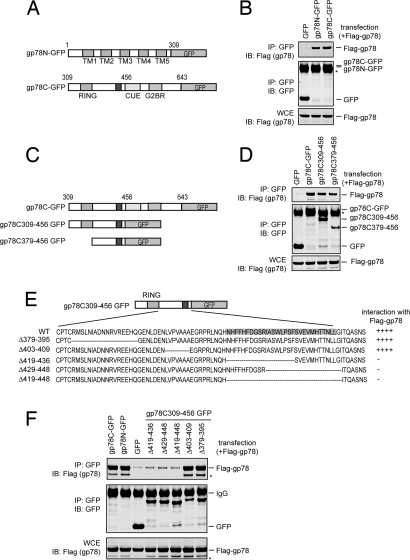
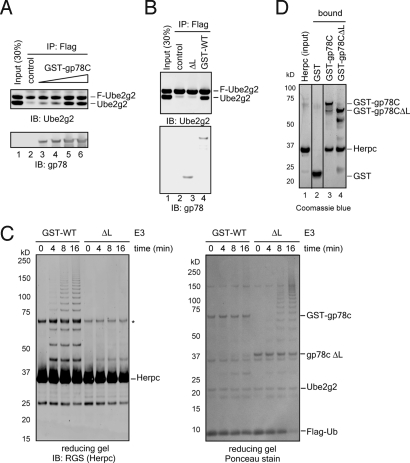
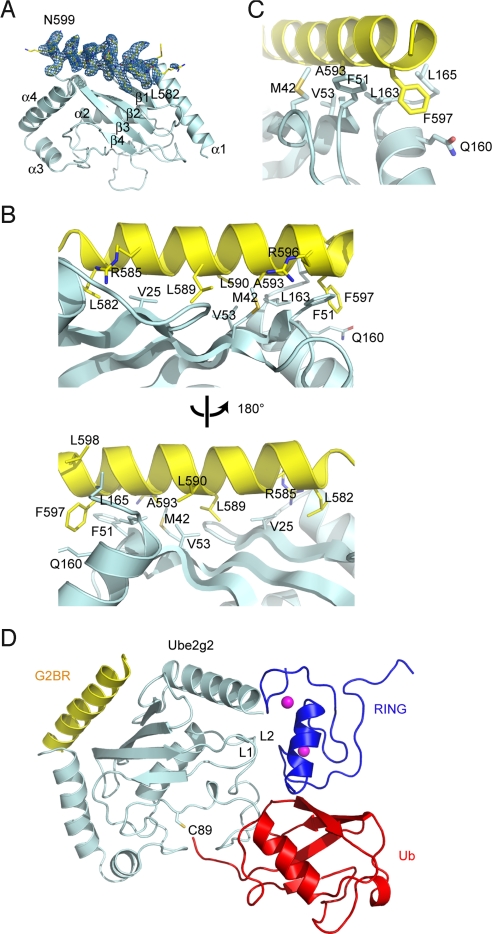
Lys-48-linked polyubiquitination regulates a variety of cellular processes by targeting ubiquitinated proteins to the proteasome for degradation. Although polyubiquitination had been presumed to occur by transferring ubiquitin molecules, one at a time, from an E2 active site to a substrate, we recently showed that the endoplasmic reticulum-associated RING finger ubiquitin ligase gp78 can mediate the preassembly of Lys-48-linked polyubiquitin chains on the catalytic cysteine of its cognate E2 Ube2g2 and subsequent transfer to a substrate. Active site-linked polyubiquitin chains are detected in cells on Ube2g2 and its yeast homolog Ubc7p, but how these chains are assembled is unclear. Here, we show that gp78 forms an oligomer via 2 oligomerization sites, one of which is a hydrophobic segment located in the gp78 cytosolic domain. We further demonstrate that a gp78 oligomer can simultaneously associate with multiple Ube2g2 molecules. This interaction is mediated by a novel Ube2g2 surface distinct from the predicted RING binding site. Our data suggest that a large gp78-Ube2g2 heterooligomer brings multiple Ube2g2 molecules into close proximity, allowing ubiquitin moieties to be transferred between neighboring Ube2g2s to form active site-linked polyubiquitin chains.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
-
- Weissman AM. Themes and variations on ubiquitylation. Nat Rev Mol Cell Biol. 2001;2:169–178. - PubMed
-
- Welchman RL, Gordon C, Mayer RJ. Ubiquitin and ubiquitin-like proteins as multifunctional signals. Nat Rev Mol Cell Biol. 2005;6:599–609. - PubMed
-
- Mukhopadhyay D, Riezman H. Proteasome-independent functions of ubiquitin in endocytosis and signaling. Science. 2007;315:201–205. - PubMed
-
- Pickart CM. Mechanisms underlying ubiquitination. Annu Rev Biochem. 2001;70:503–533. - PubMed
-
- Jin J, Li X, Gygi SP, Harper JW. Dual E1 activation systems for ubiquitin differentially regulate E2 enzyme charging. Nature. 2007;447:1135–1138. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
