

Cellular expression, trafficking, and function of two isoforms of human ULBP5/RAET1G
- PMID: 19223974
- PMCID: PMC2637608
- DOI: 10.1371/journal.pone.0004503
Cellular expression, trafficking, and function of two isoforms of human ULBP5/RAET1G
Erratum in
- PLoS ONE. 2009;4(3). doi: 10.1371/annotation/34238ee8-5128-4f1d-9ae2-6906c9764e49 doi: 10.1371/annotation/34238ee8-5128-4f1d-9ae2-6906c9764e49
- PLoS ONE. 2009;4(3). doi: 10.1371/annotation/8501cd33-6c9e-437c-9ea1-f96fbb4140d6 doi: 10.1371/annotation/8501cd33-6c9e-437c-9ea1-f96fbb4140d6
Abstract
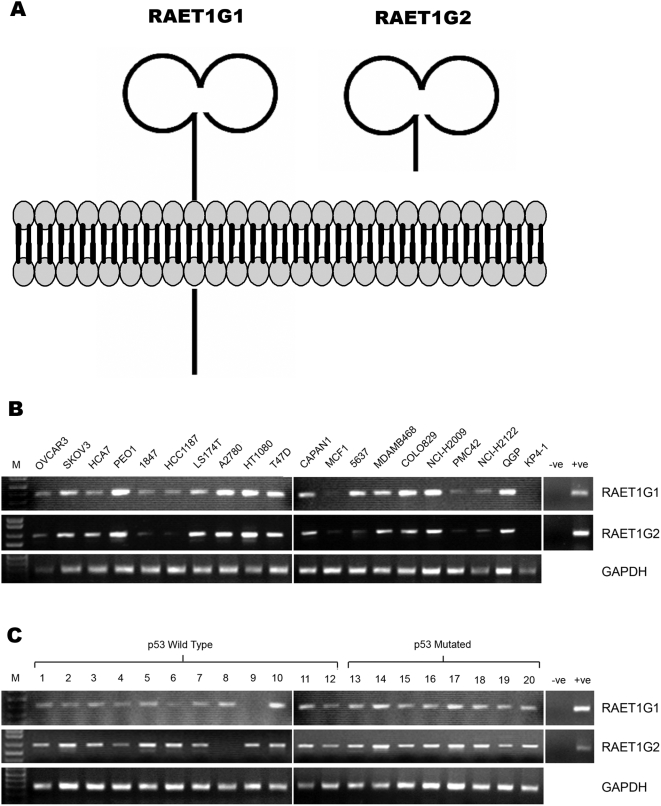
Background: The activating immunoreceptor NKG2D is expressed on Natural Killer (NK) cells and subsets of T cells. NKG2D contributes to anti-tumour and anti-viral immune responses in vitro and in vivo. The ligands for NKG2D in humans are diverse proteins of the MIC and ULBP/RAET families that are upregulated on the surface of virally infected cells and tumours. Two splicing variants of ULBP5/RAET1G have been cloned previously, but not extensively characterised.
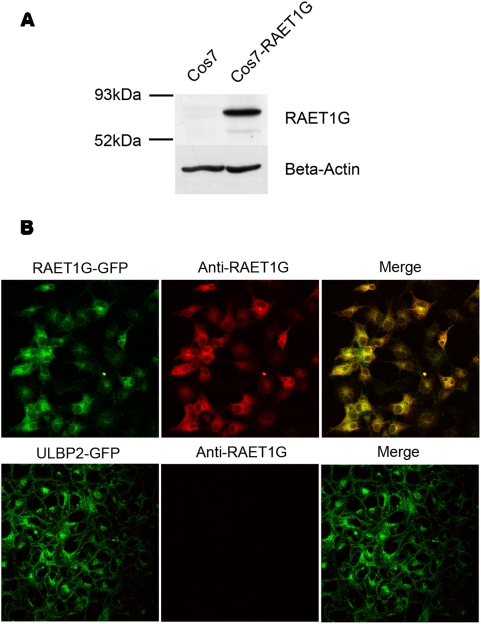
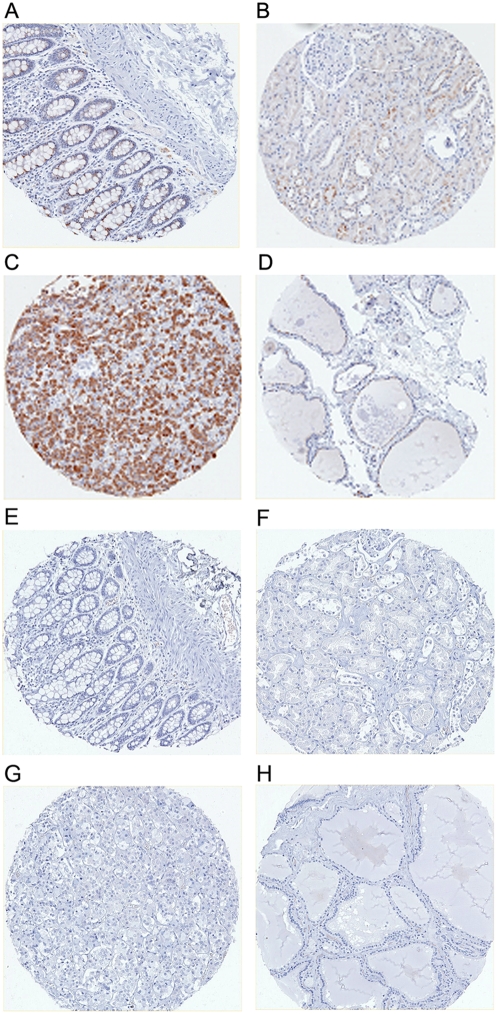
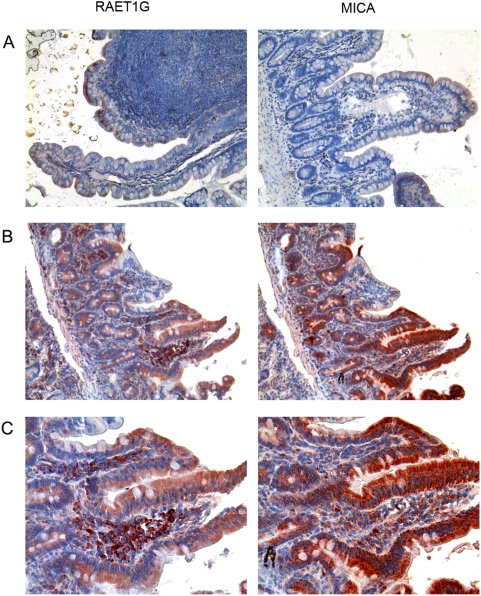
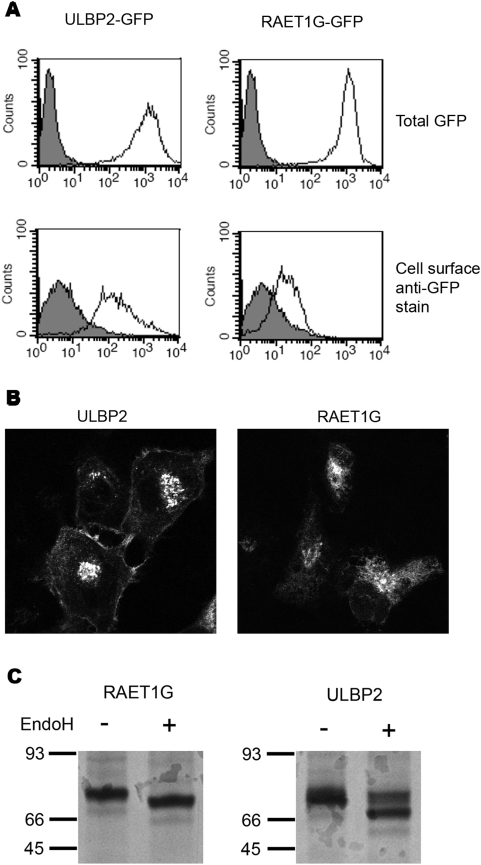
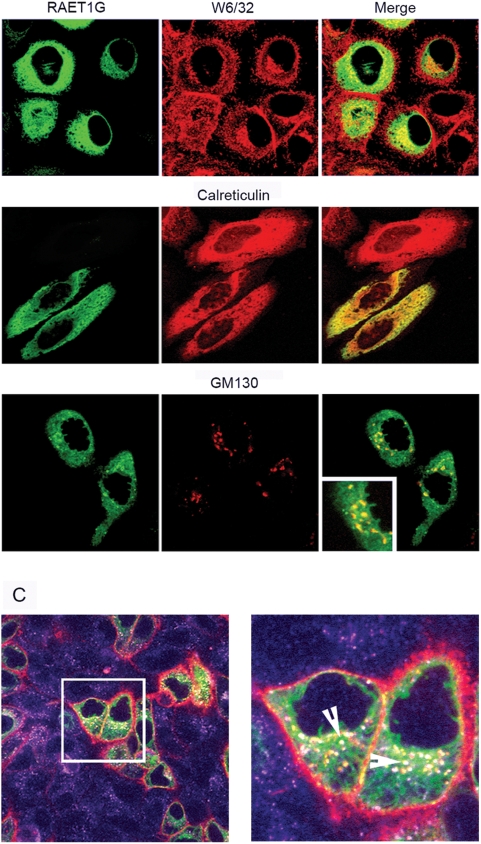
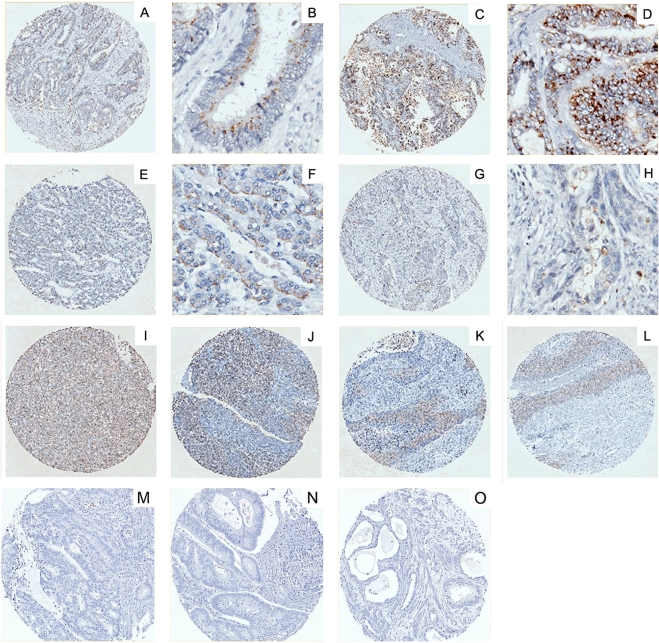
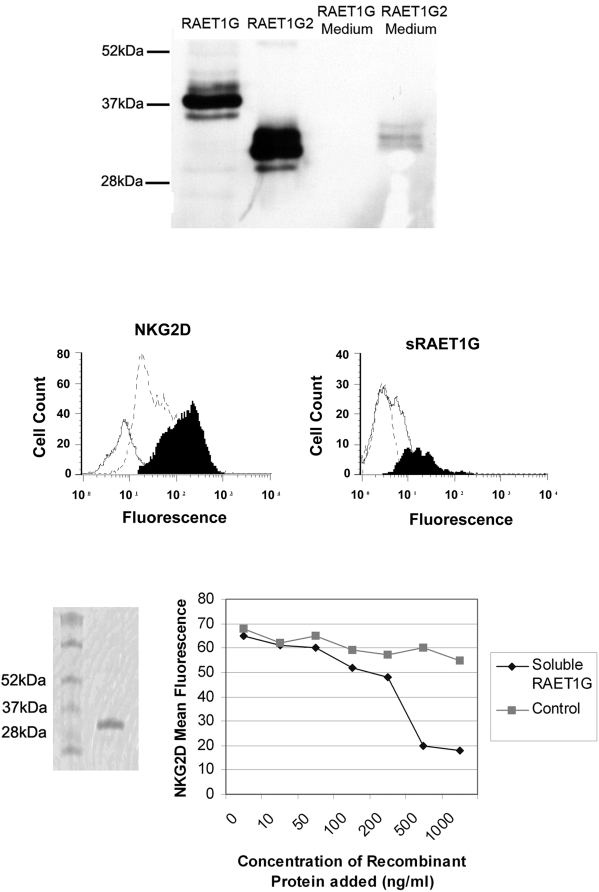
Methodology/principal findings: We pursue a number of approaches to characterise the expression, trafficking, and function of the two isoforms of ULBP5/RAET1G. We show that both transcripts are frequently expressed in cell lines derived from epithelial cancers, and in primary breast cancers. The full-length transcript, RAET1G1, is predicted to encode a molecule with transmembrane and cytoplasmic domains that are unique amongst NKG2D ligands. Using specific anti-RAET1G1 antiserum to stain tissue microarrays we show that RAET1G1 expression is highly restricted in normal tissues. RAET1G1 was expressed at a low level in normal gastrointestinal epithelial cells in a similar pattern to MICA. Both RAET1G1 and MICA showed increased expression in the gut of patients with celiac disease. In contrast to healthy tissues the RAET1G1 antiserum stained a wide variety or different primary tumour sections. Both endogenously expressed and transfected RAET1G1 was mainly found inside the cell, with a minority of the protein reaching the cell surface. Conversely the truncated splicing variant of RAET1G2 was shown to encode a soluble molecule that could be secreted from cells. Secreted RAET1G2 was shown to downregulate NKG2D receptor expression on NK cells and hence may represent a novel tumour immune evasion strategy.
Conclusions/significance: We demonstrate that the expression patterns of ULBP5RAET1G are very similar to the well-characterised NKG2D ligand, MICA. However the two isoforms of ULBP5/RAET1G have very different cellular localisations that are likely to reflect unique functionality.
Conflict of interest statement
Figures
References
-
- Raulet DH. Roles of the NKG2D immunoreceptor and its ligands. Nat Rev Immunol. 2003;3:781–790. - PubMed
-
- Groh V, Smythe K, Dai Z, Spies T. Fas-ligand-mediated paracrine T cell regulation by the receptor NKG2D in tumor immunity. Nat Immunol. 2006;7:755–762. - PubMed
-
- Saez-Borderias A, Guma M, Angulo A, Bellosillo B, Pende D, et al. Expression and function of NKG2D in CD4+ T cells specific for human cytomegalovirus. Eur J Immunol. 2006;36:3198–3206. - PubMed
-
- Gonzalez S, Groh V, Spies T. Immunobiology of human NKG2D and its ligands. Curr Top Microbiol Immunol. 2006;298:121–138. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
