

Experimental infection of pigs with the human 1918 pandemic influenza virus
- PMID: 19224986
- PMCID: PMC2668479
- DOI: 10.1128/JVI.02399-08
Experimental infection of pigs with the human 1918 pandemic influenza virus
Abstract
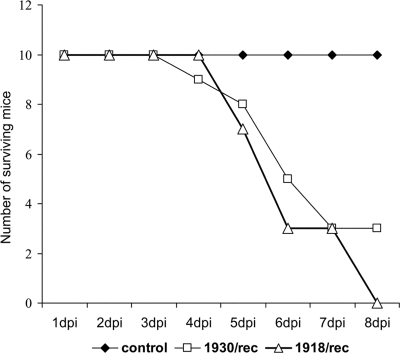
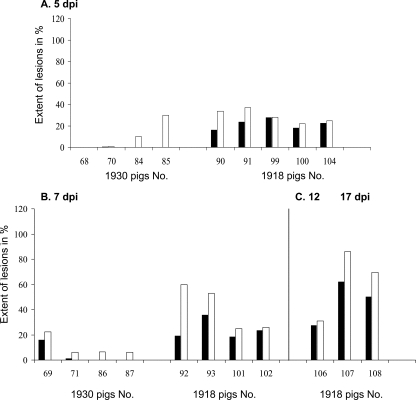
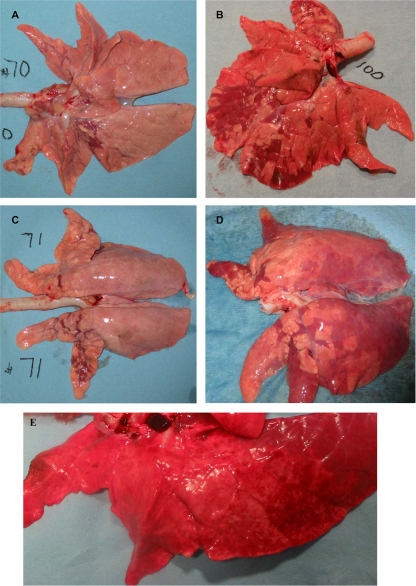
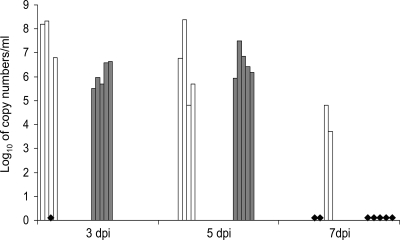
Swine influenza was first recognized as a disease entity during the 1918 "Spanish flu" pandemic. The aim of this work was to determine the virulence of a plasmid-derived human 1918 pandemic H1N1 influenza virus (reconstructed 1918, or 1918/rec, virus) in swine using a plasmid-derived A/swine/Iowa/15/1930 H1N1 virus (1930/rec virus), representing the first isolated influenza virus, as a reference. Four-week-old piglets were inoculated intratracheally with either the 1930/rec or the 1918/rec virus or intranasally with the 1918/rec virus. A transient increase in temperature and mild respiratory signs developed postinoculation in all virus-inoculated groups. In contrast to other mammalian hosts (mice, ferrets, and macaques) where infection with the 1918/rec virus was lethal, the pigs did not develop severe respiratory distress or become moribund. Virus titers in the lower respiratory tract as well as macro- and microscopic lesions at 3 and 5 days postinfection (dpi) were comparable between the 1930/rec and 1918/rec virus-inoculated animals. In contrast to the 1930/rec virus-infected animals, at 7 dpi prominent lung lesions were present in only the 1918/rec virus-infected animals, and all the piglets developed antibodies at 7 dpi. Presented data support the hypothesis that the 1918 pandemic influenza virus was able to infect and replicate in swine, causing a respiratory disease, and that the virus was likely introduced into the pig population during the 1918 pandemic, resulting in the current lineage of the classical H1N1 swine influenza viruses.
Figures
Similar articles
-
Experimental infection with a Thai reassortant swine influenza virus of pandemic H1N1 origin induced disease.Virol J. 2013 Mar 16;10:88. doi: 10.1186/1743-422X-10-88. Virol J. 2013. PMID: 23497073 Free PMC article.
-
Pathogenesis of swine influenza virus (Thai isolates) in weanling pigs: an experimental trial.Virol J. 2009 Mar 25;6:34. doi: 10.1186/1743-422X-6-34. Virol J. 2009. PMID: 19317918 Free PMC article.
-
Antigenically Diverse Swine Origin H1N1 Variant Influenza Viruses Exhibit Differential Ferret Pathogenesis and Transmission Phenotypes.J Virol. 2018 May 14;92(11):e00095-18. doi: 10.1128/JVI.00095-18. Print 2018 Jun 1. J Virol. 2018. PMID: 29540597 Free PMC article.
-
History of Swine influenza viruses in Asia.Curr Top Microbiol Immunol. 2013;370:57-68. doi: 10.1007/82_2011_179. Curr Top Microbiol Immunol. 2013. PMID: 21948002 Review.
-
[Swine influenza virus: evolution mechanism and epidemic characterization--a review].Wei Sheng Wu Xue Bao. 2009 Sep;49(9):1138-45. Wei Sheng Wu Xue Bao. 2009. PMID: 20030049 Review. Chinese.
Cited by
-
Efficacy of influenza vaccination and tamiflu® treatment--comparative studies with Eurasian Swine influenza viruses in pigs.PLoS One. 2013 Apr 22;8(4):e61597. doi: 10.1371/journal.pone.0061597. Print 2013. PLoS One. 2013. PMID: 23630601 Free PMC article.
-
Animal models for the risk assessment of viral pandemic potential.Lab Anim Res. 2020 Apr 22;36:11. doi: 10.1186/s42826-020-00040-6. eCollection 2020. Lab Anim Res. 2020. PMID: 32337177 Free PMC article. Review.
-
Protective efficacy of a broadly cross-reactive swine influenza DNA vaccine encoding M2e, cytotoxic T lymphocyte epitope and consensus H3 hemagglutinin.Virol J. 2012 Jun 27;9:127. doi: 10.1186/1743-422X-9-127. Virol J. 2012. PMID: 22738661 Free PMC article.
-
Systems approaches to influenza-virus host interactions and the pathogenesis of highly virulent and pandemic viruses.Semin Immunol. 2013 Oct 31;25(3):228-39. doi: 10.1016/j.smim.2012.11.001. Epub 2012 Dec 5. Semin Immunol. 2013. PMID: 23218769 Free PMC article. Review.
-
Cross talk between animal and human influenza viruses.Annu Rev Anim Biosci. 2013 Jan;1:21-42. doi: 10.1146/annurev-animal-031412-103733. Epub 2013 Jan 1. Annu Rev Anim Biosci. 2013. PMID: 25387011 Free PMC article. Review.
References
-
- Bikour, M. H., E. H. Frost, S. Deslandes, B. Talbot, and Y. Elazhary. 1995. Persistence of a 1930 swine influenza A (H1N1) virus in Quebec. J. Gen. Virol. 762539-2547. - PubMed
-
- Brown, I. H. 2000. The epidemiology and evolution of influenza viruses in pigs. Vet. Microbiol. 7429-46. - PubMed
-
- Choi, Y. K., T. D. Nguyen, H. Ozaki, R. J. Webby, P. Puthavathana, C. Buranathal, A. Chaisingh, P. Auewarakul, N. T. Hanh, S. K. Ma, P. Y. Hui, Y. Guan, J. S. Peiris, and R. G. Webster. 2005. Studies of H5N1 influenza virus infection of pigs by using viruses isolated in Vietnam and Thailand in 2004. J. Virol. 7910821-10825. - PMC - PubMed
-
- Easterday, B. C. 2003. Swine influenza: historical perspective, p. 241-244. In P. Martelli, S. Caviarani, and A. Lavazza (ed.), Proceedings of the 4th International Symposium on Emerging and Re-emerging Pig Diseases, Rome, 29 to 2 July 2003. University of Parma, Parma, Italy.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
