

Intersubunit interactions modulate pH-induced activation of membrane fusion by the Junin virus envelope glycoprotein GPC
- PMID: 19224989
- PMCID: PMC2668491
- DOI: 10.1128/JVI.02410-08
Intersubunit interactions modulate pH-induced activation of membrane fusion by the Junin virus envelope glycoprotein GPC
Abstract
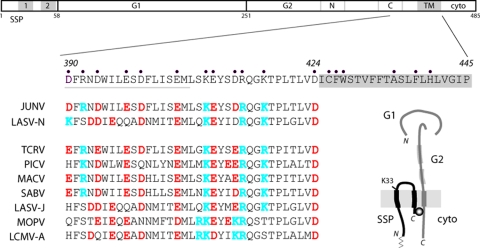
The mature arenavirus envelope glycoprotein GPC is a tripartite complex comprising a stable signal peptide (SSP) in addition to the receptor-binding (G1) and transmembrane fusion (G2) subunits. We have shown previously that SSP is a key element in GPC-mediated membrane fusion, and that GPC sensitivity to acidic pH is modulated in part through the lysine residue at position 33 in the ectodomain loop of SSP (J. York and J. H. Nunberg, J. Virol. 80:7775-7780, 2006). A glutamine substitution at this position stabilizes the native GPC complex and thereby prevents the induction of pH-dependent membrane fusion. In efforts to identify the intersubunit interactions of K33, we performed alanine-scanning mutagenesis at charged residues in the membrane-proximal ectodomain of G2 and determined the ability of these mutations to rescue the fusion deficiency in K33Q GPC. Four second-site mutations that specifically complement K33Q were identified (D400A, E410A, R414A, and K417A). Moreover, complementation was also observed at three hydrophobic positions in the membrane-spanning domain of G2 (F427, W428, and F438). Interestingly, all of the complementing mutations restored wild-type pH sensitivity to the K33Q mutant, while none themselves affected the pH of membrane fusion. Our studies demonstrate a specific interaction between SSP and G2 that is involved in priming the native GPC complex for pH-induced membrane fusion. Importantly, this pH-dependent interaction has been shown to be vulnerable to small-molecule compounds that stabilize the native complex and prevent the activation of membrane fusion. A detailed mechanistic understanding of the control of GPC-mediated membrane fusion will be important in guiding the development of effective therapeutics against arenaviral hemorrhagic fever.
Figures



Similar articles
-
Epistastic Interactions within the Junín Virus Envelope Glycoprotein Complex Provide an Evolutionary Barrier to Reversion in the Live-Attenuated Candid#1 Vaccine.J Virol. 2017 Dec 14;92(1):e01682-17. doi: 10.1128/JVI.01682-17. Print 2018 Jan 1. J Virol. 2017. PMID: 29070682 Free PMC article.
-
A novel zinc-binding domain is essential for formation of the functional Junín virus envelope glycoprotein complex.J Virol. 2007 Dec;81(24):13385-91. doi: 10.1128/JVI.01785-07. Epub 2007 Oct 10. J Virol. 2007. PMID: 17928348 Free PMC article.
-
Myristoylation of the Arenavirus Envelope Glycoprotein Stable Signal Peptide Is Critical for Membrane Fusion but Dispensable for Virion Morphogenesis.J Virol. 2016 Aug 26;90(18):8341-50. doi: 10.1128/JVI.01124-16. Print 2016 Sep 15. J Virol. 2016. PMID: 27412594 Free PMC article.
-
The curious case of arenavirus entry, and its inhibition.Viruses. 2012 Jan;4(1):83-101. doi: 10.3390/v4010083. Epub 2012 Jan 13. Viruses. 2012. PMID: 22355453 Free PMC article. Review.
-
Structure-function relationship of the mammarenavirus envelope glycoprotein.Virol Sin. 2016 Oct;31(5):380-394. doi: 10.1007/s12250-016-3815-4. Epub 2016 Aug 4. Virol Sin. 2016. PMID: 27562602 Free PMC article. Review.
Cited by
-
Envelope glycoprotein of arenaviruses.Viruses. 2012 Oct 17;4(10):2162-81. doi: 10.3390/v4102162. Viruses. 2012. PMID: 23202458 Free PMC article. Review.
-
An antibody directed against the fusion peptide of Junin virus envelope glycoprotein GPC inhibits pH-induced membrane fusion.J Virol. 2010 Jun;84(12):6119-29. doi: 10.1128/JVI.02700-09. Epub 2010 Apr 14. J Virol. 2010. PMID: 20392854 Free PMC article.
-
Identification of Clotrimazole Derivatives as Specific Inhibitors of Arenavirus Fusion.J Virol. 2019 Mar 5;93(6):e01744-18. doi: 10.1128/JVI.01744-18. Print 2019 Mar 15. J Virol. 2019. PMID: 30626681 Free PMC article.
-
Restoration of virulence in the attenuated Candid#1 vaccine virus requires reversion at both positions 168 and 427 in the envelope glycoprotein GPC.J Virol. 2024 Apr 16;98(4):e0011224. doi: 10.1128/jvi.00112-24. Epub 2024 Mar 20. J Virol. 2024. PMID: 38506509 Free PMC article.
-
Strategies of rational and structure-driven vaccine design for Arenaviruses.Infect Genet Evol. 2024 Sep;123:105626. doi: 10.1016/j.meegid.2024.105626. Epub 2024 Jun 20. Infect Genet Evol. 2024. PMID: 38908736 Free PMC article. Review.
References
-
- Bolken, T. C., S. Laquerre, Y. Zhang, T. R. Bailey, D. C. Pevear, S. S. Kickner, L. E. Sperzel, K. F. Jones, T. K. Warren, S. Amanda Lund, D. L. Kirkwood-Watts, D. S. King, A. C. Shurtleff, M. C. Guttieri, Y. Deng, M. Bleam, and D. E. Hruby. 2006. Identification and characterization of potent small molecule inhibitor of hemorrhagic fever New World arenaviruses. Antivir. Res. 6986-89. - PMC - PubMed
-
- Borrow, P., and M. B. A. Oldstone. 1994. Mechanism of lymphocytic choriomeningitis virus entry into cells. Virology 1981-9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
