

Feature selectivity of the gamma-band of the local field potential in primate primary visual cortex
- PMID: 19225593
- PMCID: PMC2622750
- DOI: 10.3389/neuro.01.037.2008
Feature selectivity of the gamma-band of the local field potential in primate primary visual cortex
Abstract
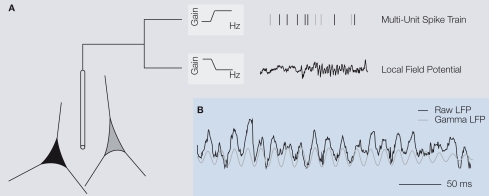
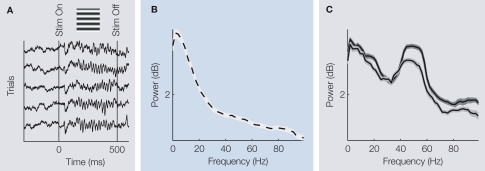
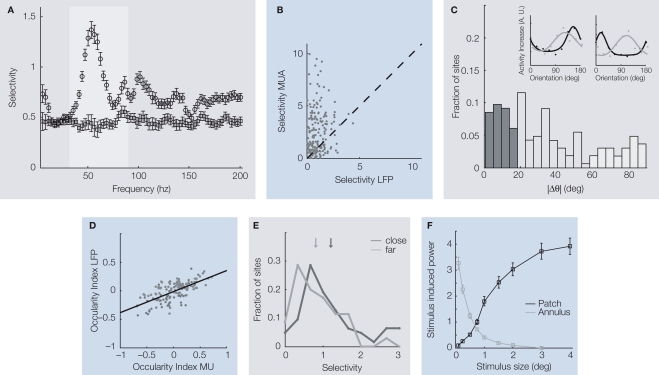
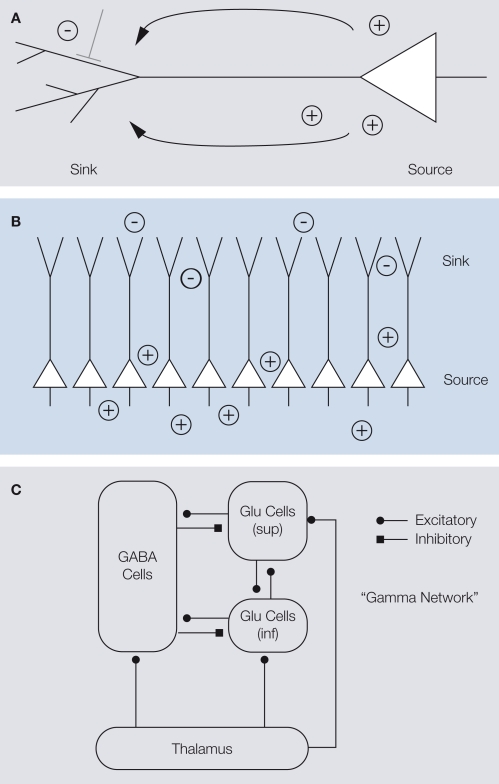
Extracellular voltage fluctuations (local field potentials, LFPs) reflecting neural mass action are ubiquitous across species and brain regions. Numerous studies have characterized the properties of LFP signals in the cortex to study sensory and motor computations as well as cognitive processes like attention, perception and memory. In addition, its extracranial counterpart - the electroencephalogram - is widely used in clinical applications. However, the link between LFP signals and the underlying activity of local populations of neurons remains largely elusive. Here, we review recent work elucidating the relationship between spiking activity of local neural populations and LFP signals. We focus on oscillations in the gamma-band (30-90 Hz) of the LFP in the primary visual cortex (V1) of the macaque that dominate during visual stimulation. Given that in area V1 much is known about the properties of single neurons and the cortical architecture, it provides an excellent opportunity to study the mechanisms underlying the generation of the LFP.
Keywords: feature selectivity; gamma-band; local field potential; primary visual cortex; spatial resolution.
Figures
References
Grants and funding
LinkOut - more resources
Full Text Sources