

Review
doi: 10.1021/cr8004197.
Intramembrane proteolysis
Affiliations
- PMID: 19226105
- PMCID: PMC2667872
- DOI: 10.1021/cr8004197
Item in Clipboard
Review
Intramembrane proteolysis
Chem Rev.
2009 Apr.
No abstract available
Figures
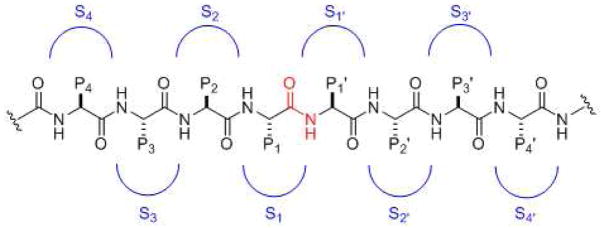
General substrate and substrate binding sites for proteases. The scissile amide functionality is in red. Moving left from the cleavage site are residues P1, P2, P3, P4, etc. Moving from the right of the cleavage site are residues P1′, P2′, P3′, P4′, etc. Corresponding binding pockets on the protease for these residues are S1, S2, S3, S4, etc. on the left side and S1′, S2′, S3′, S4′, etc. on the right side.
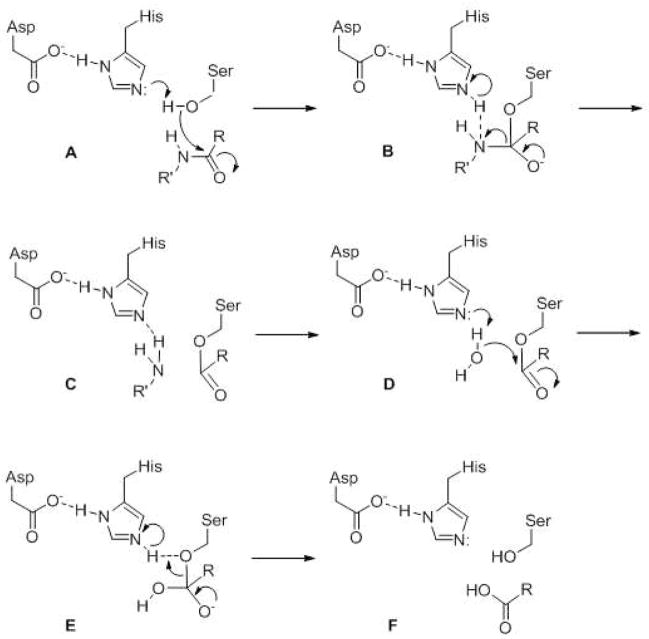
General catalytic mechanism for serine proteases. Active site serine, histidine and (often) aspartate comprise a catalytic triad that work together first to activate the serine hydroxyl group for nucleophilic attack of the amide carbonyl. After release of the amino group, water is then activated for nucleophilic attack of the ester carbonyl of the protease-substrate covalent intermediate. The carboxylate is then released, and the enzyme is back in its original state and ready for another round of catalysis.
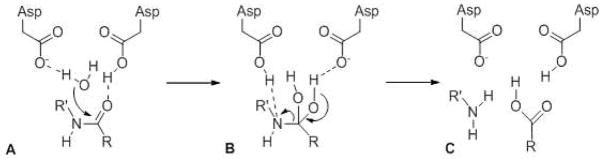
General catalytic mechanism for aspartyl proteases. Two active site aspartates activate water and the scissile amide bond of the substrate via hydrogen bonding. Nucleophilic attack of water on the amide carbonyl forms a tetrahedral gem-diol intermediate that fragments to the two cleavage products.
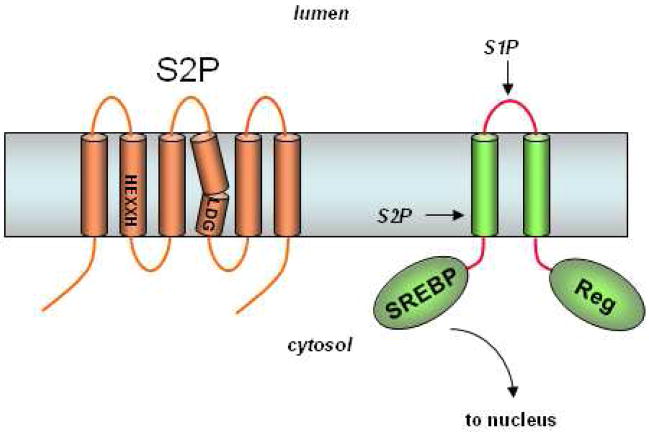
S2P contains conserved HEXXH and LDG motifs found in metalloproteases. SREBP is first cleaved by S1P in the luminal loop. The regulatory domain (Reg) interacts with the cholesterol-sensing SCAP to ensure that S1P proteolysis only occurs when cholesterol levels are low. Subsequent intramembrane proteolysis releases this transcription factor for expression of genes essential to cholesterol and fatty acid synthesis.
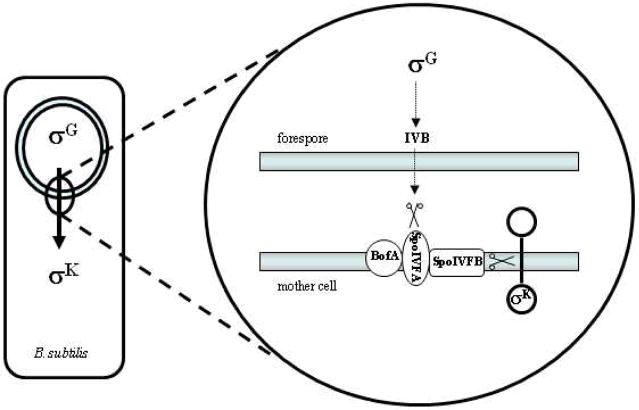
Bacillus subtilisS2P-like protease SpoIVFB and sporulation. Upon engulfment of the forespore by the mother cell, a signaling pathway involving the transcription factor σG is initiated in the forespore that triggers the synthesis of the IVB serine protease. This protease degrades SpoIVFA, which along with BofA serves to inhibit SpoIVFB. With the inhibition of the S2P-like protease released, SpoIVFB cleaves pro-σK, allowing this transcription factor to signal in the mother cell for further factors needed for spore maturation.
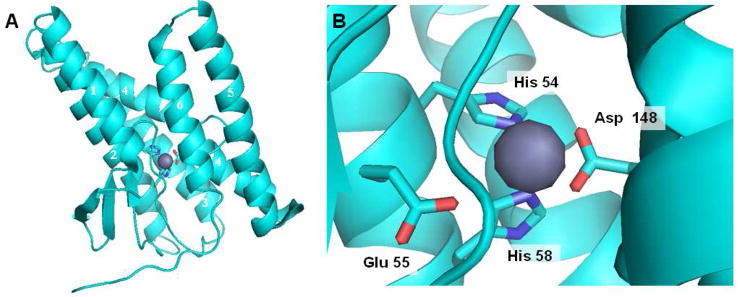
Structure of archaeal MjS2P. (A) The six transmembrane domains (TMDs) are arranged as a helical bundle, with two histidines from TMD 2 and an aspartate from TMD 4 coordinated to the zinc atom (grey). A second crystallized conformation has TMD 1 and TMD 6 much further apart, suggesting that this is the site of lateral gating by which the transmembrane of the substrate accesses the internal active site. (B) Close up of the active site, explicitly showing another residue (a glutamate in TMD 2) coordinated with the zinc. This glutamate activates water for catalysis.
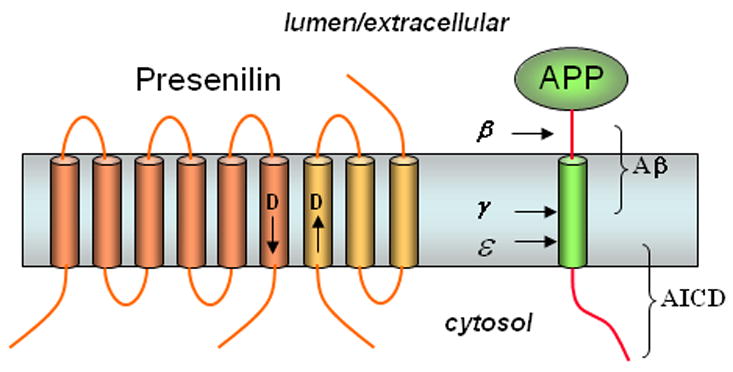
Presenilin, the γ-secretase complex, and the proteolysis of APP to Aβ. Presenilin is processes into two pieces, an N-terminal fragment (NTF, dark portion) and a C-terminal fragment (CTF, light portion) that remain associated. Each fragment donates one aspartate essential for γ-secretase activity (arrows near these aspartates denote N- to C-terminal directionality of the protein sequence3). APP is first cleaved in the extracellular domain by β-secretase, and the remnant is cleaved twice within the membrane by γ-secretase to produce the Aβ peptide of Alzheimer’s disease (secreted) and the intracellular domain (AICD, freed into the cytosol).
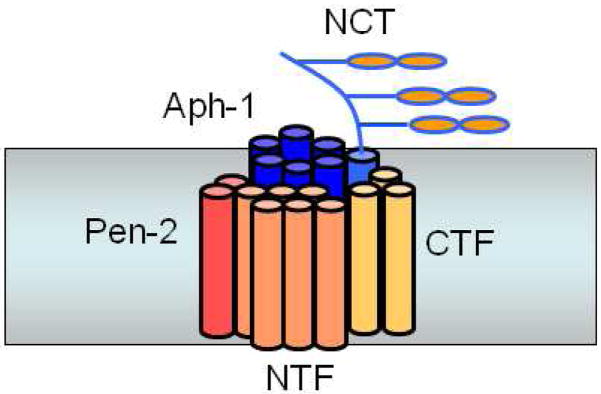
Presenilin interacts with three other membrane proteins, nicastrin, Aph-1, and Pen-2, whereupon presenilin is cleaved into NTF and CTF to form active γ-secretase.
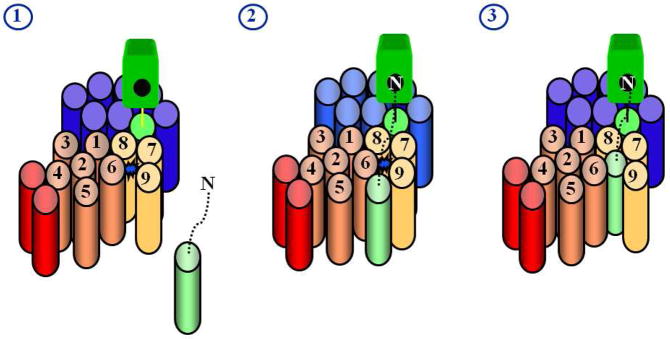
Model for how substrates interact with presenilin. Substrate is light green, presenilin NTF is copper, presenilin CTF is gold, Pen-2 is red, Aph-1 is blue, and nicastrin is bright green. The active site (represented by the blue star in stage 1), containing water and two aspartates, is thought to be sequestered away from the hydrophobic environment of the lipid bilayer, necessitating an external docking site for the transmembrane domain of the substrate. Studies with chemical probes (helical peptides and transition-state analogues) support this model (see text). The transmembrane domain of the substrate interacts with the docking site, while the N-terminus of the substrate putatively interacts with the Nicastrin ectodomain (stage 2). The transmembrane domain of the substrate then passes either in whole or in part into the active site for proteolysis (stage 3).
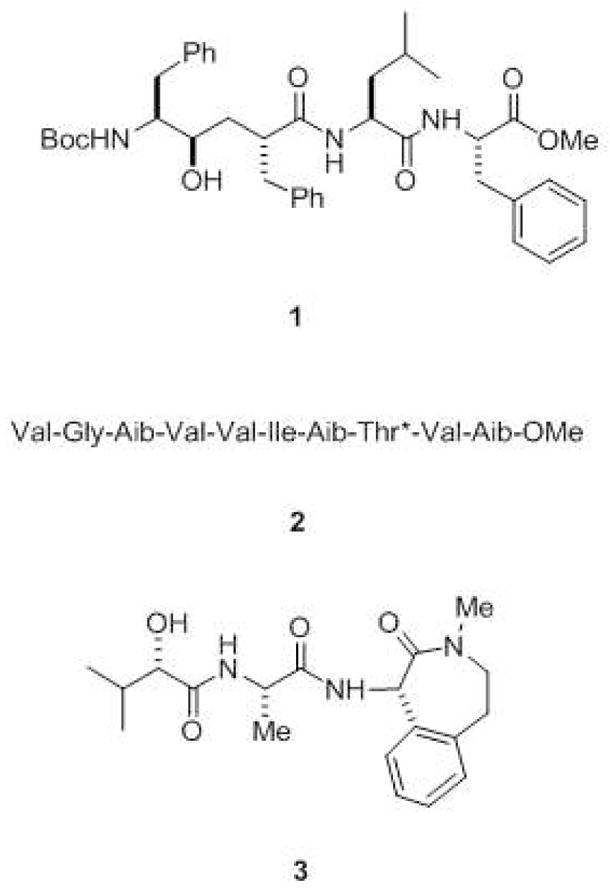
Inhibitors of γ-secretase. Transition-state analogue inhibitors such as 1 include hydroxyl-containing moieties that interact with the catalytic aspartates of aspartyl proteases. Helical peptide inhibitors include α-aminoisobutyric acid (Aib)-containing substrate mimics such as 2 (*denotes that the threonine residue contains an O-benzyl group). These helical peptides mimic the APP transmembrane domain and interact with the substrate docking site on the protease. Also shown is the potent benzodiazepine inhibitor 3 (LY-450,139), which is in late-stage clinical trials for the treatment of Alzheimer’s disease.
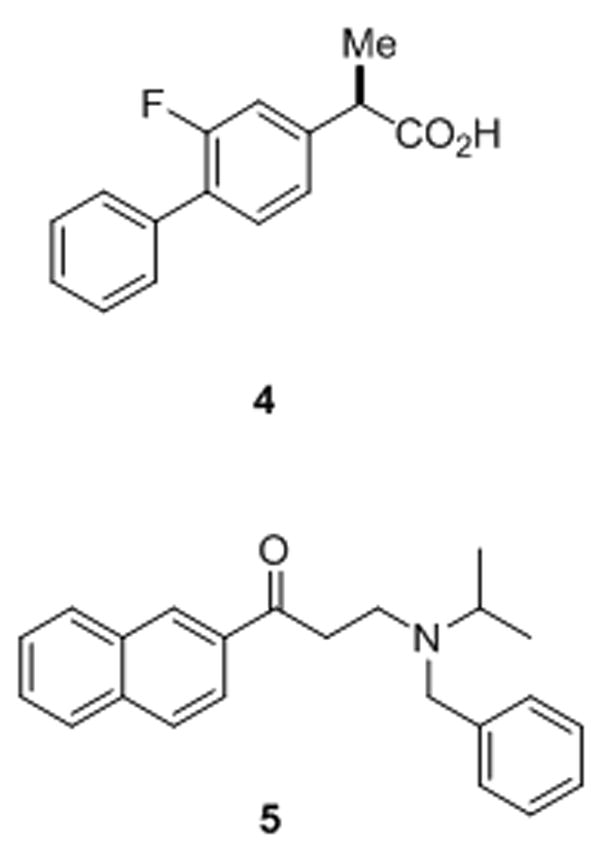
Modulators of γ-secretase. NSAID-like modulator 4 (R-Flurbiprofen or tarenflurbil), which is in late-stage clinical trials for Alzheimer’s disease, shifts where γ-secretase cuts APP, reducing the aggregation-prone Aβ42 and elevating more soluble Aβ38. In contrast, naphthyl ketone 5 inhibits total Aβ production without interfering with the ability of γ-secretase to cleave Notch receptor substrates.
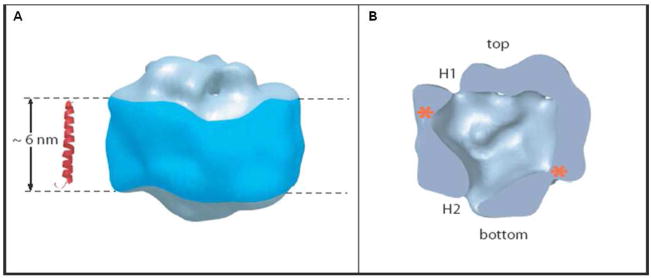
Structure of the γ-secretase complex as deduced by electron microscopy. (A) Although the resolution is relatively low (~15 Ǻ) and the structure is globular, (B) a central cavity of low density suggests the location of the active site, and two ports, H1 and H2, suggest site of water entry. Reprinted with permission from reference . Copyright 2006 National Academy of Science.
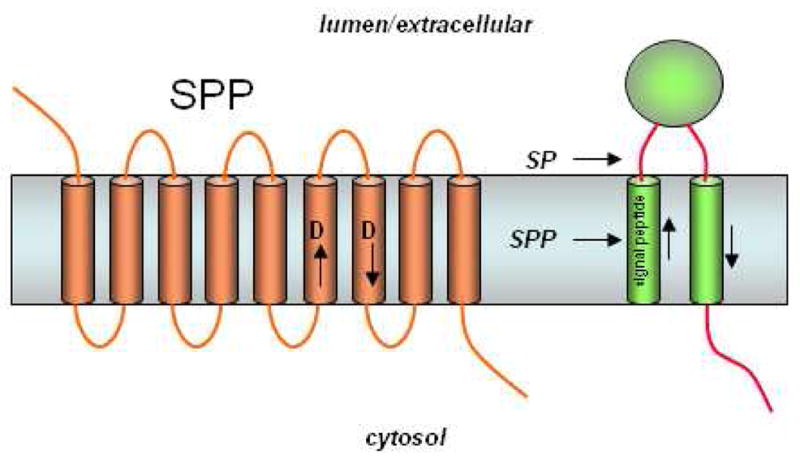
The presenilin homolog signal peptide peptidase (SPP). Signal peptides are removed from membrane proteins via signal peptidase (SP), and these peptides are released from the membrane by SPP-mediated intramembrane proteolysis. SPP, like presenilin, contains two aspartates essential for protease activity, but the conserved aspartate-containing motifs are in the opposite orientation compared with their presenilin counterparts (cf. Fig. 7). Consistent with the flipped orientation of SPP vis-à-vis presenilin, the substrates of these two proteases also run in the opposite direction. Unlike presenilin, SPP apparently does not require other protein cofactors or cleavage into two subunits for proteolytic activity.
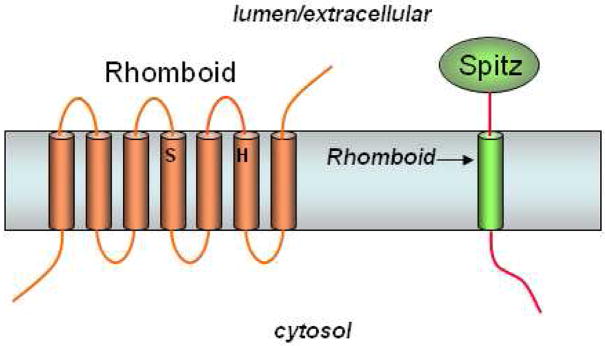
Rhomboid serine proteases. Rhomboids contain a conserved serine and histidine, which comprise a putative catalytic dyad of a serine protease. Rhomboid-1 cleaves within the transmembrane region of the Drosophila EGF-like growth factor Spitz.
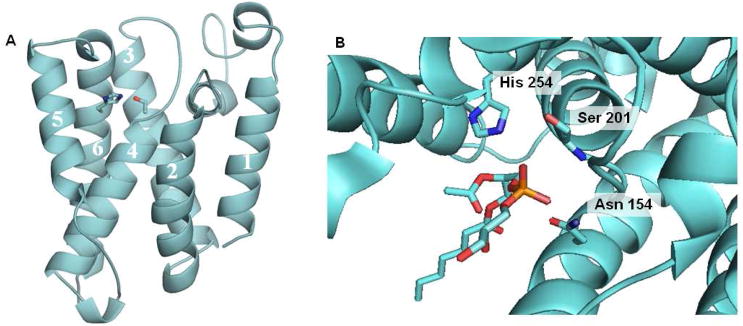
Structure of E. coli Rhomboid GlpG. (A) The serine in transmembrane domain 4 and the histidine in transmembrane domain 6 are coordinated in a manner consistent with known serine proteases and at a depth within the membrane consistent with the site of proteolysis of Rhomboid substrates. (B) Close up view of the active site from a crystal structure with a bound phospholipid. The interaction of the phosphate group with the backbone NH of serine 102 and with the side chain of asparagines 154 suggests the site of the oxyanion hole.
References
-
- Barrett AJ, Rawlings ND, Woessner JF, editors. The Handbook of Proteolytic Enzymes. Academic Press, Ltd.; Amsterdam: 2004.
-
- Turk B. Nat Rev Drug Discov. 2006;5:785. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
