

IL-32-dependent effects of IL-1beta on endothelial cell functions
- PMID: 19228941
- PMCID: PMC2656174
- DOI: 10.1073/pnas.0813334106
IL-32-dependent effects of IL-1beta on endothelial cell functions
Abstract
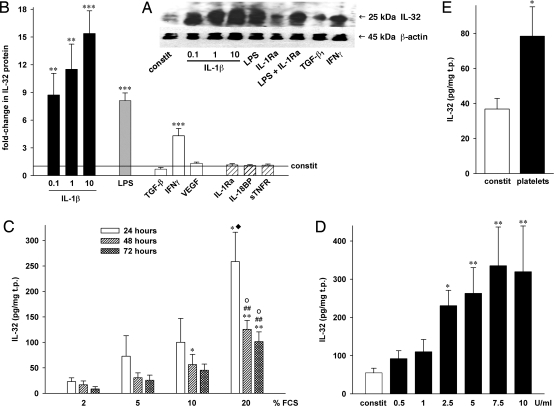
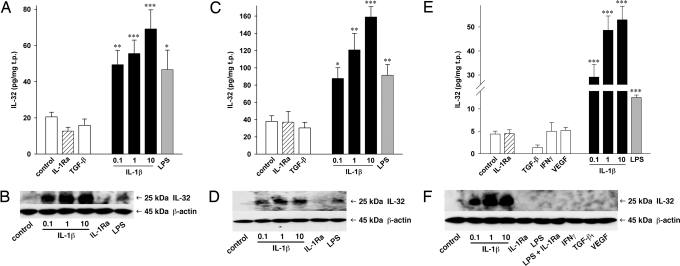
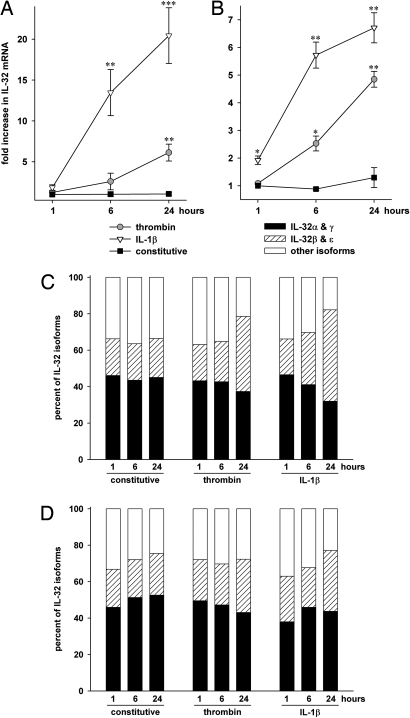
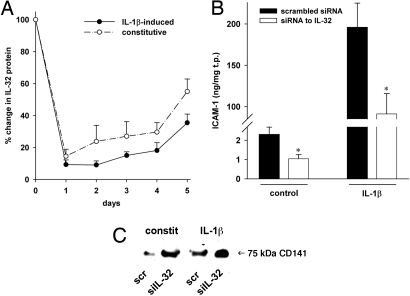
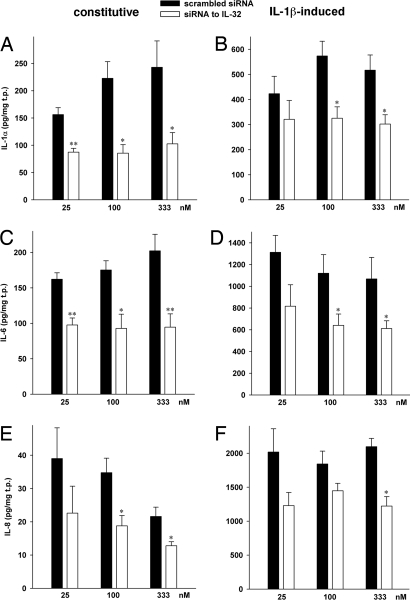
Increasing evidence demonstrates that interleukin (IL)-32 is a pro-inflammatory cytokine, inducing IL-1alpha, IL-1beta, IL-6, tumor necrosis factor (TNF)-alpha, and chemokines via nuclear factor (NF)-kappaB, p38 mitogen-activated protein kinase (MAPK), and activating protein (AP)-1 activation. Here we report that IL-32 is expressed and is also functional in human vascular endothelial cells (EC) of various origins. Compared with primary blood monocytes, high levels of IL-32 are constitutively produced in human umbilical vein EC (HUVEC), aortic macrovascular EC, and cardiac as well as pulmonary microvascular EC. At concentrations as low as 0.1 ng/ml, IL-1beta stimulated IL-32 up to 15-fold over constitutive levels, whereas 10 ng/ml of TNFalpha or 100 ng/ml of lipopolysaccharide (LPS) were required to induce similar quantities of IL-32. IL-1beta-induced IL-32 was reduced by inhibition of the IkappaB kinase-beta/NF-kappaB and ERK pathways. In addition to IL-1beta, pro-coagulant concentrations of thrombin or fresh platelets increased IL-32 protein up to 6-fold. IL-1beta and thrombin induced an isoform-switch in steady-state mRNA levels from IL-32alpha/gamma to beta/epsilon. Adult EC responded in a similar fashion. To prove functionality, we silenced endogenous IL-32 with siRNA, decreasing intracellular IL-32 protein levels by 86%. The knockdown of IL-32 resulted in reduction of constitutive as well as IL-1beta-induced intercellular adhesion molecule-1 (ICAM-1) (of 55% and 54%, respectively), IL-1alpha (of 62% and 43%), IL-6 (of 53% and 43%), and IL-8 (of 46% and 42%). In contrast, the anti-inflammatory/anti-coagulant CD141/thrombomodulin increased markedly when IL-32 was silenced. This study introduces IL-32 as a critical regulator of endothelial function, expanding the properties of this cytokine relevant to coagulation, endothelial inflammation, and atherosclerosis.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kim SH, et al. Interleukin-32: A cytokine and inducer of TNFalpha. Immunity. 2005;22:131–142. - PubMed
-
- Nold MF, et al. Endogenous IL-32 controls cytokine and HIV-1 production. J Immunol. 2008;181:557–565. - PubMed
-
- Goda C, et al. Involvement of IL-32 in activation-induced cell death in T cells. Int Immunol. 2006;18:233–240. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
