

Involvement of the cytoplasmic C-terminal domain of connexin43 in neuronal migration
- PMID: 19228955
- PMCID: PMC6666339
- DOI: 10.1523/JNEUROSCI.5025-08.2009
Involvement of the cytoplasmic C-terminal domain of connexin43 in neuronal migration
Abstract
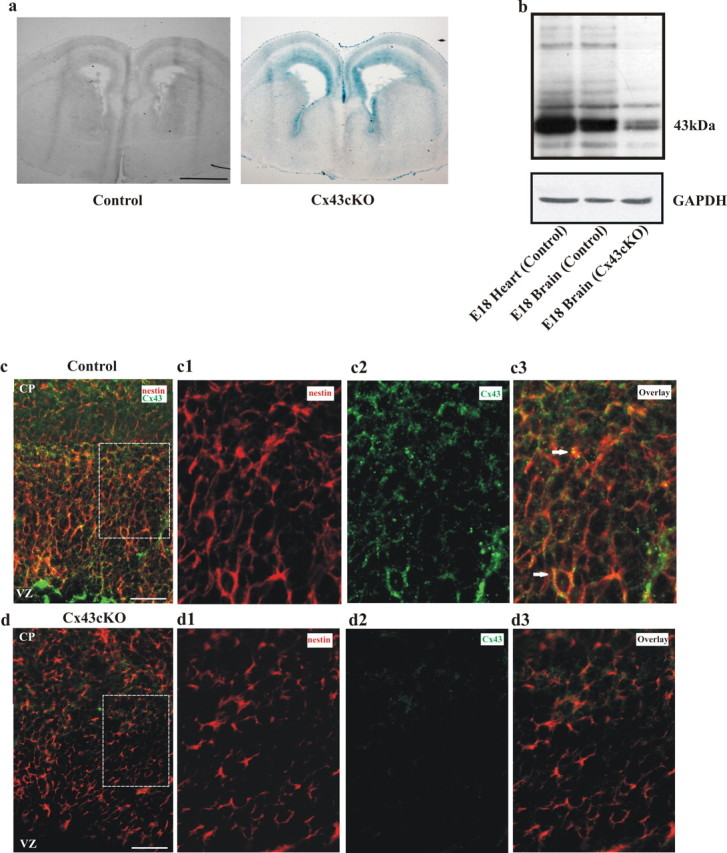
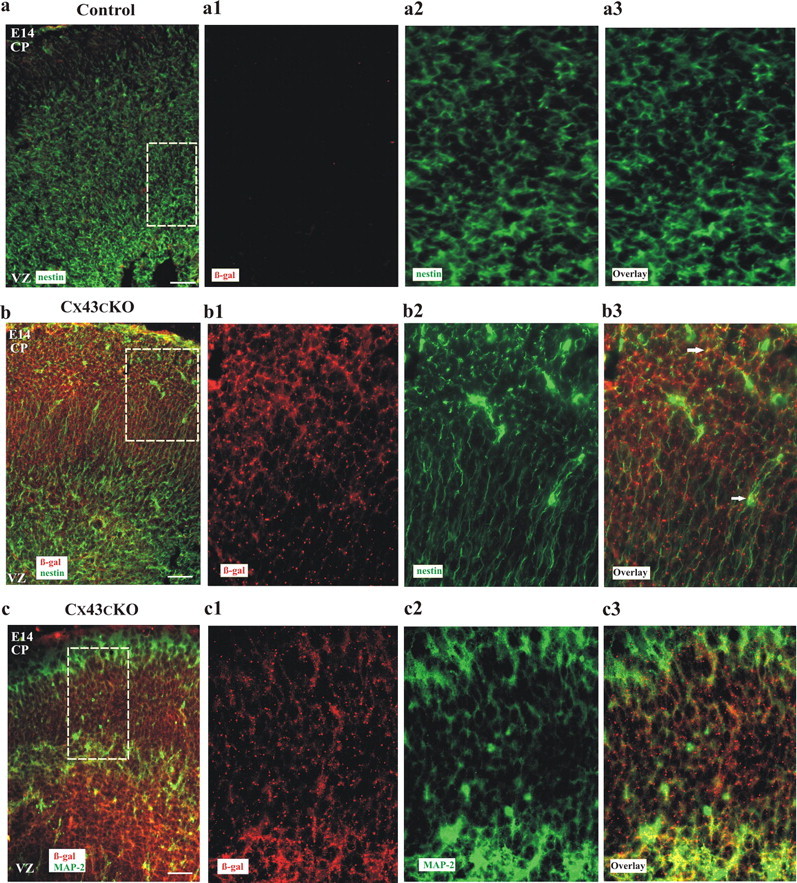
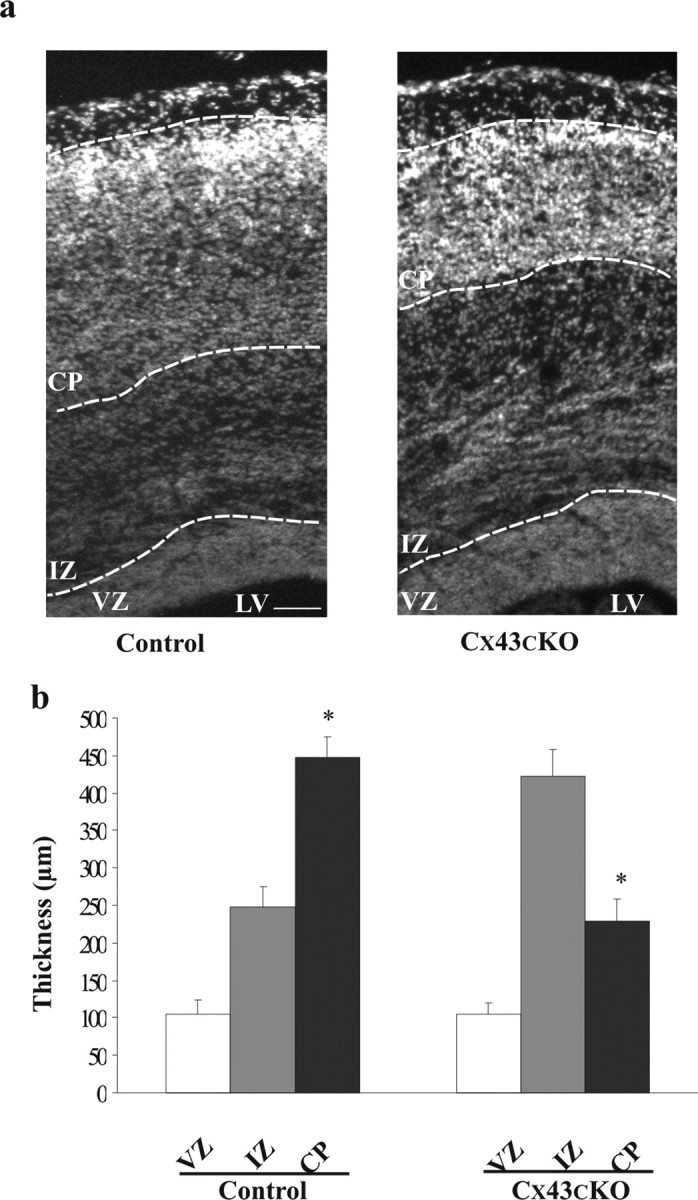
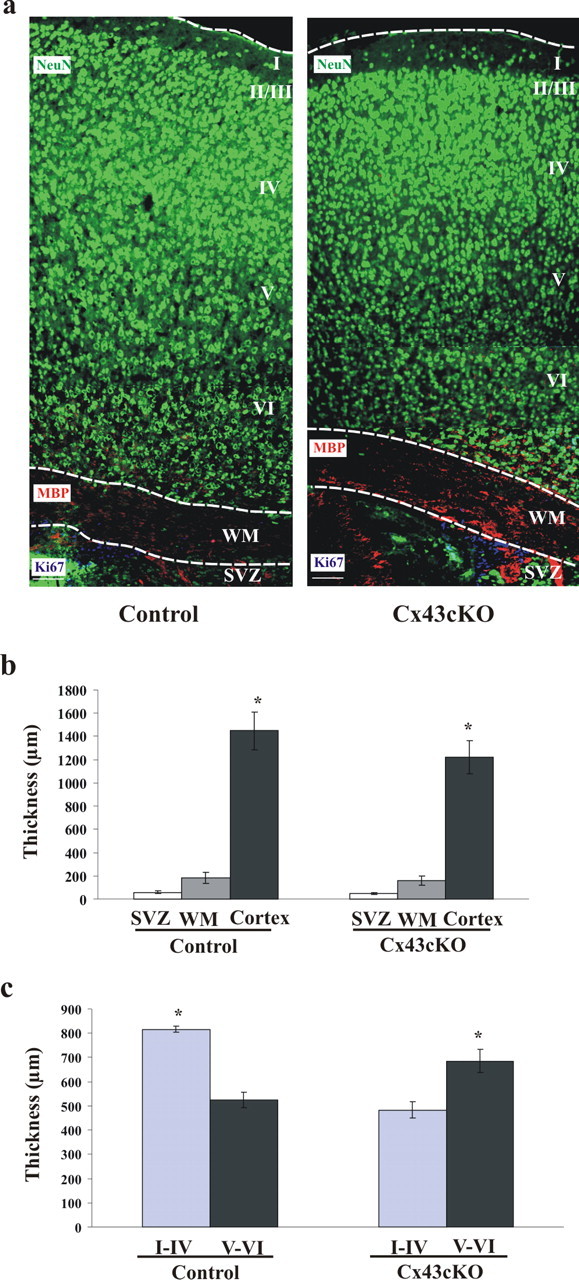
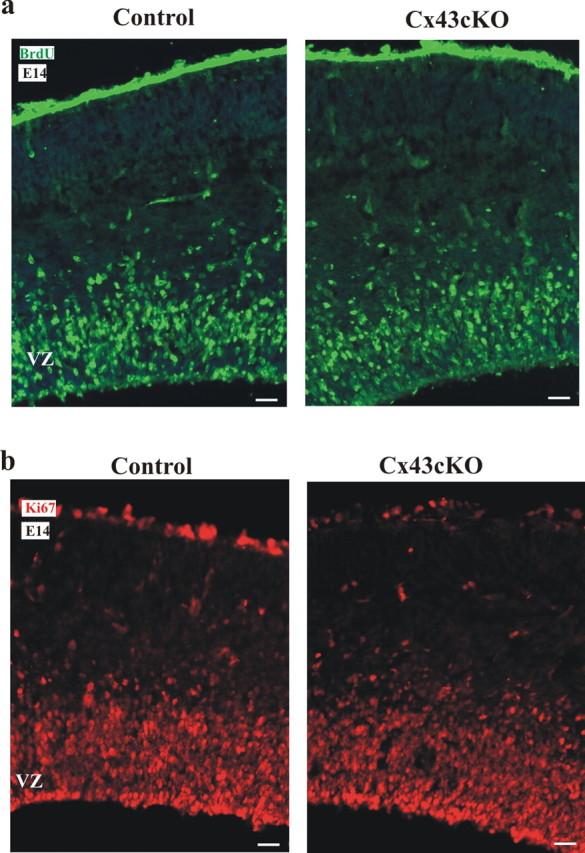
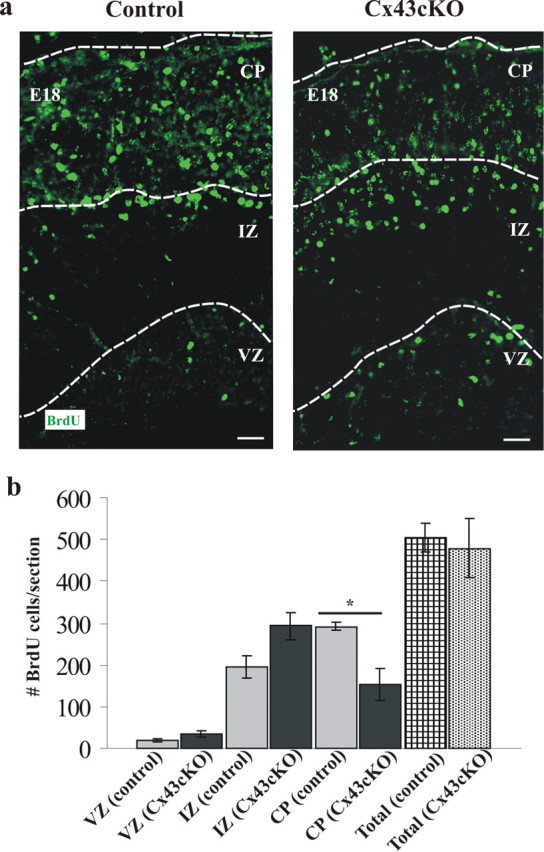
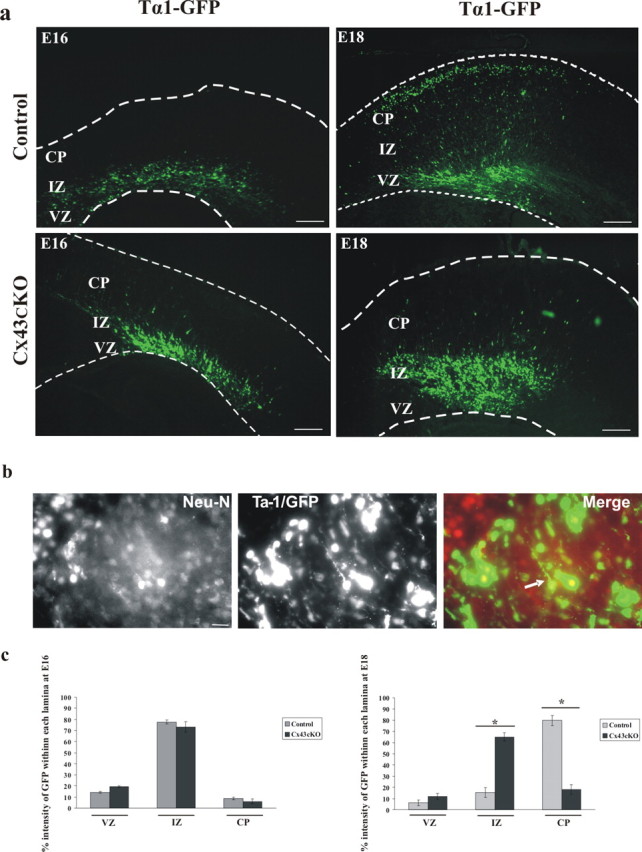
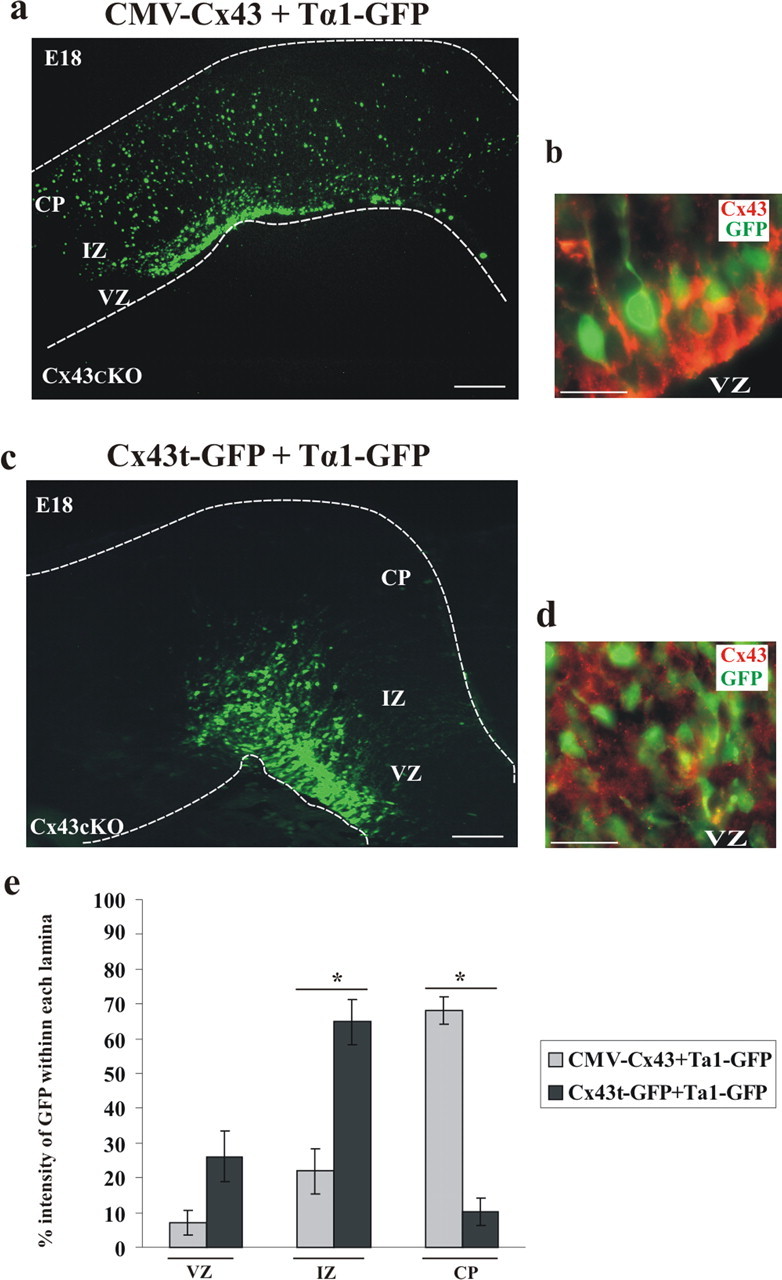
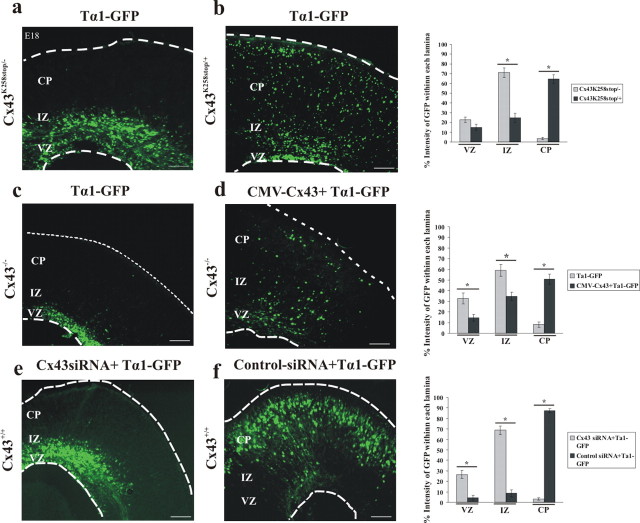
During brain development, young neurons closely associate with radial glial while migrating from the ventricular zone (VZ) to the cortical plate (CP) of the neocortex. It has been shown previously that gap junctions are needed for this migration to occur properly, but the precise mechanism responsible is still in question. Here, we used Cre recombinase, driven by the nestin promoter, to conditionally knock-out a floxed coding DNA of the connexin43 (Cx43) gene in mice. Radial glia in the VZ normally express connexin43. They undergo divisions that produce neurons and astrocytes and serve as migratory guides for the daughter cells that they produce. Based on histological analysis, we suggest that removing Cx43 from radial glia alters the normal lamination of the mouse neocortex. To monitor newborn neurons during development, we introduced a plasmid containing green fluorescent protein driven by a neuronal (Talpha1 tubulin) promoter into the embryonic neocortex using in utero electroporation. The transfected migrating neurons remain in the VZ/intermediate zone (IZ) of the Cx43 conditional knock-out (Cx43cKO) animals, whereas in Cx43(fl/fl) mice, neurons migrate through the IZ into the CP, indicating that deletion of Cx43 from nestin-positive cells disrupts neuronal migration. We were able to rescue migration of Cx43cKO neurons by electroporating a cytomegalovirus-Cx43 expression plasmid into the embryonic cortex. In contrast, a C-terminal truncated form of Cx43 failed to rescue neuronal migration. In addition, Cx43(K258stop) mice, in which Cx43 lacks the last 125 amino acid residues of the cytoplasmic C-terminal domain, gave results similar to those seen with the Cx43cKO mice. This study illustrates that deletion of the C-terminal domain of Cx43 alters neuronal migration in the neocortex.
Figures
References
-
- Anthony TE, Klein C, Fishell G, Heintz N. Radial glia serve as neuronal progenitors in all regions of the central nervous system. Neuron. 2004;41:881–890. - PubMed
-
- Barnabé-Heider F, Wasylnka JA, Fernandes KJ, Porsche C, Sendtner M, Kaplan DR, Miller FD. Evidence that embryonic neurons regulate the onset of cortical gliogenesis via cardiotrophin-1. Neuron. 2005;48:253–265. - PubMed
-
- Bates DC, Sin WC, Aftab Q, Naus CC. Connexin43 enhances glioma invasion by a mechanism involving the carboxy terminus. Glia. 2007;55:1554–1564. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous