

Correlates of perceptual learning in an oculomotor decision variable
- PMID: 19228966
- PMCID: PMC2668818
- DOI: 10.1523/JNEUROSCI.3962-08.2009
Correlates of perceptual learning in an oculomotor decision variable
Abstract
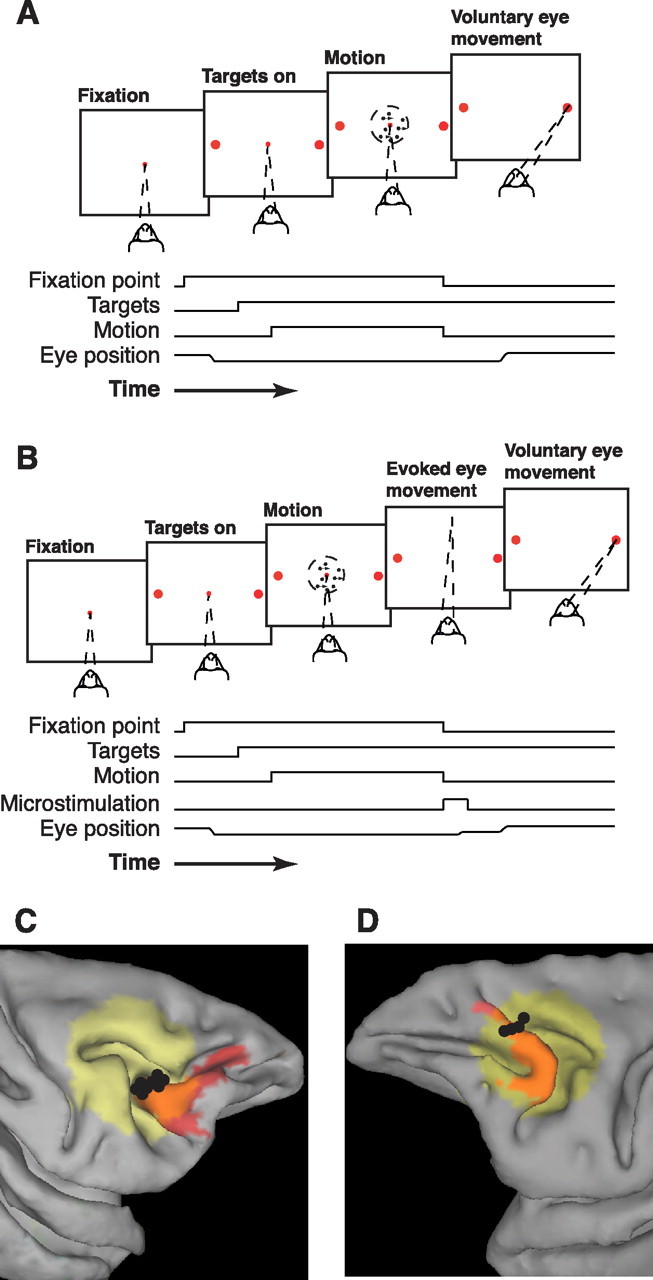
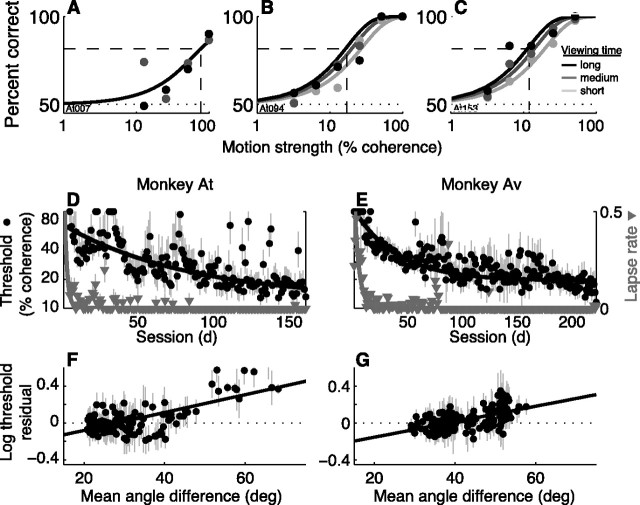
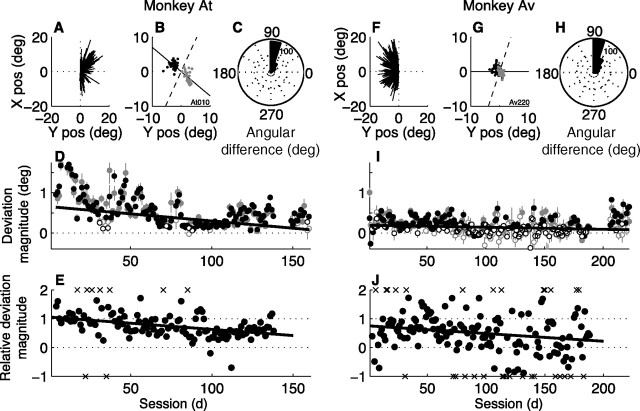
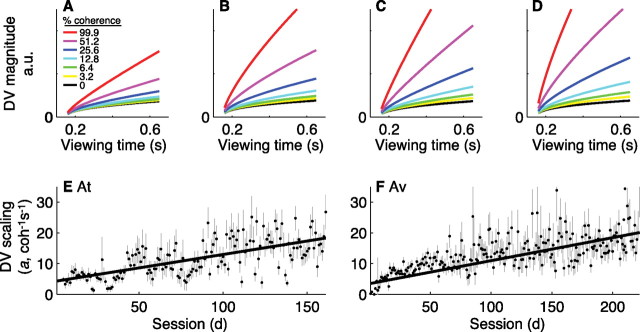
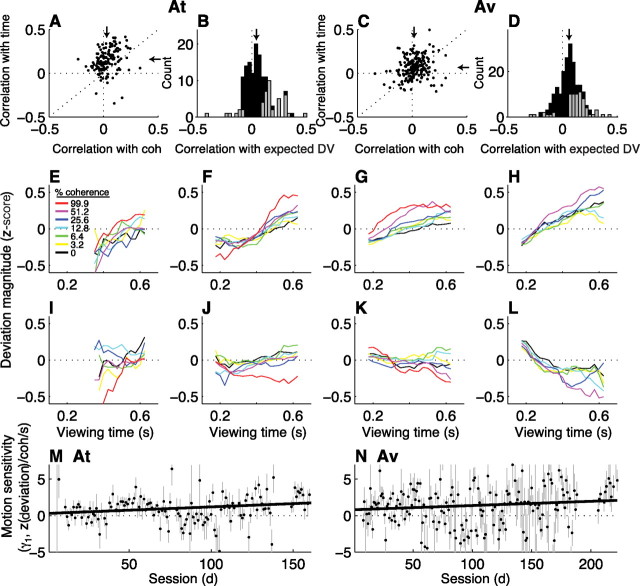
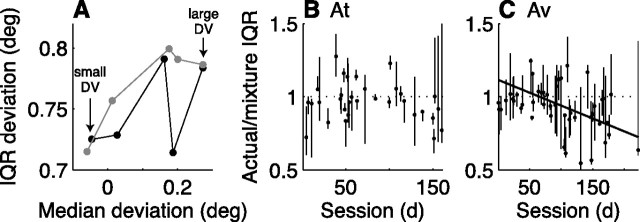
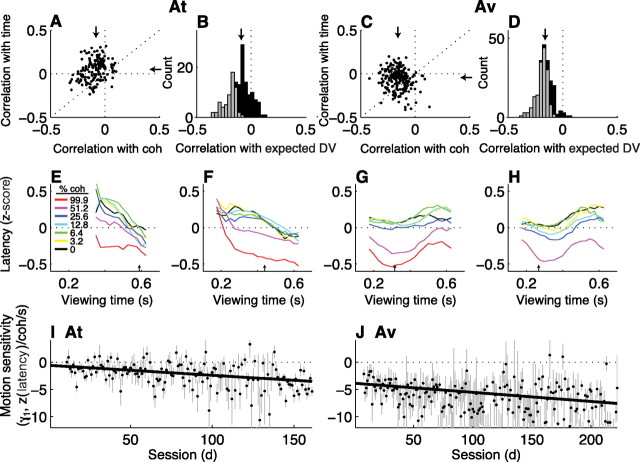
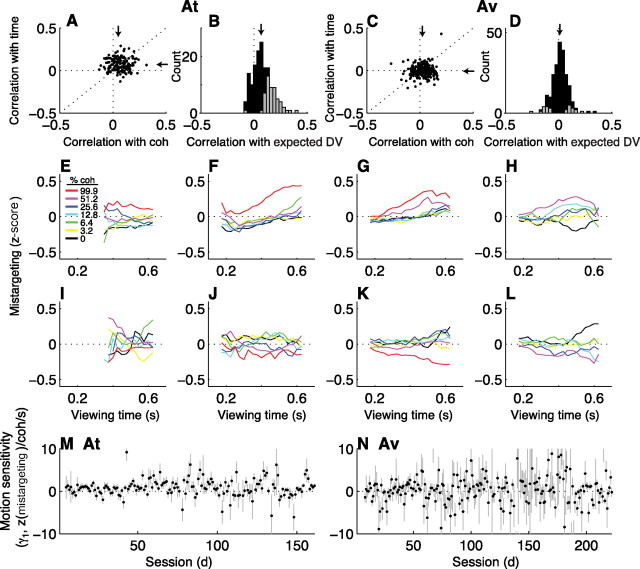
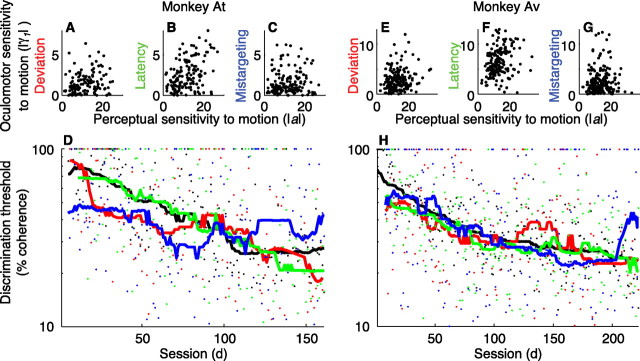
In subjects trained extensively to indicate a perceptual decision with an action, neural commands that generate the action can represent the process of forming the decision. However, it is unknown whether this representation requires overtraining or reflects a more general link between perceptual and motor processing. We examined how perceptual processing is represented in motor commands in naive monkeys being trained on a demanding perceptual task, as they first establish the sensory-motor association and then learn to form more accurate perceptual judgments. The task required the monkeys to decide the direction of random-dot motion and respond with an eye movement to one of two visual targets. Using electrically evoked saccades, we examined oculomotor commands that developed during motion viewing. Throughout training, these commands tended to reflect both the subsequent binary choice of saccade target and the weighing of graded motion evidence used to arrive at that choice. Moreover, these decision-related oculomotor signals, along with the time needed to initiate the voluntary saccadic response, changed steadily as training progressed, approximately matching concomitant improvements in behavioral sensitivity to the motion stimulus. Thus, motor circuits may have general access to perceptual processing used to select between actions, even without extensive training. The results also suggest a novel candidate mechanism for some forms of perceptual learning, in which the brain learns rapidly to treat a perceptual decision as a problem of action selection and then over time to use sensory input more effectively to guide the selection process.
Figures
Similar articles
-
The relative influences of priors and sensory evidence on an oculomotor decision variable during perceptual learning.J Neurophysiol. 2008 Nov;100(5):2653-68. doi: 10.1152/jn.90629.2008. Epub 2008 Aug 27. J Neurophysiol. 2008. PMID: 18753326 Free PMC article.
-
Representation of a perceptual decision in developing oculomotor commands.Nature. 2000 Mar 23;404(6776):390-4. doi: 10.1038/35006062. Nature. 2000. PMID: 10746726
-
The influence of behavioral context on the representation of a perceptual decision in developing oculomotor commands.J Neurosci. 2003 Jan 15;23(2):632-51. doi: 10.1523/JNEUROSCI.23-02-00632.2003. J Neurosci. 2003. PMID: 12533623 Free PMC article.
-
[Parietal Association Area and Motion Information Processing].Brain Nerve. 2016 Nov;68(11):1335-1343. doi: 10.11477/mf.1416200597. Brain Nerve. 2016. PMID: 27852024 Review. Japanese.
-
Visual learning for perceptual and categorical decisions in the human brain.Vision Res. 2010 Feb 22;50(4):433-40. doi: 10.1016/j.visres.2009.09.025. Epub 2009 Oct 7. Vision Res. 2010. PMID: 19818361 Review.
Cited by
-
Biased associative representations in parietal cortex.Neuron. 2013 Jan 9;77(1):180-91. doi: 10.1016/j.neuron.2012.11.014. Neuron. 2013. PMID: 23312525 Free PMC article.
-
A low-frequency oscillatory neural signal in humans encodes a developing decision variable.Neuroimage. 2013 Dec;83:795-808. doi: 10.1016/j.neuroimage.2013.06.085. Epub 2013 Jul 18. Neuroimage. 2013. PMID: 23872495 Free PMC article.
-
Reinforcement learning can account for associative and perceptual learning on a visual-decision task.Nat Neurosci. 2009 May;12(5):655-63. doi: 10.1038/nn.2304. Epub 2009 Apr 19. Nat Neurosci. 2009. PMID: 19377473 Free PMC article.
-
Action Planning and the Timescale of Evidence Accumulation.PLoS One. 2015 Jun 12;10(6):e0129473. doi: 10.1371/journal.pone.0129473. eCollection 2015. PLoS One. 2015. PMID: 26068458 Free PMC article.
-
Relationships between the threshold and slope of psychometric and neurometric functions during perceptual learning: implications for neuronal pooling.J Neurophysiol. 2010 Jan;103(1):140-54. doi: 10.1152/jn.00744.2009. Epub 2009 Oct 28. J Neurophysiol. 2010. PMID: 19864439 Free PMC article.
References
-
- Audley RJ, Pike AR. Some alternative stochastic models of choice. Br J Math Stat Psychol. 1965;18:207–225.
-
- Ball K, Sekuler R. A specific and enduring improvement in visual motion discrimination. Science. 1982;218:697–698. - PubMed
-
- Ball K, Sekuler R. Direction-specific improvement in motion discrimination. Vision Res. 1987;27:953–965. - PubMed
-
- Bogacz R, Brown E, Moehlis J, Holmes P, Cohen JD. The physics of optimal decision making: a formal analysis of models of performance in two-alternative forced choice tasks. Psychol Rev. 2006;113:700–765. - PubMed
-
- Brainard DH. The psychophysics toolbox. Spat Vis. 1997;10:433–436. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources