

Mechanistic insights into nitrite-induced cardioprotection using an integrated metabolomic/proteomic approach
- PMID: 19229060
- PMCID: PMC6731772
- DOI: 10.1161/CIRCRESAHA.108.187005
Mechanistic insights into nitrite-induced cardioprotection using an integrated metabolomic/proteomic approach
Abstract
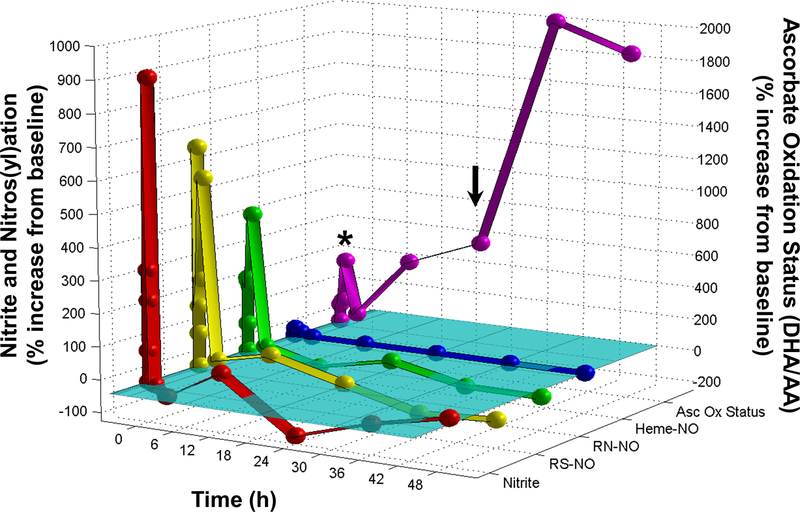
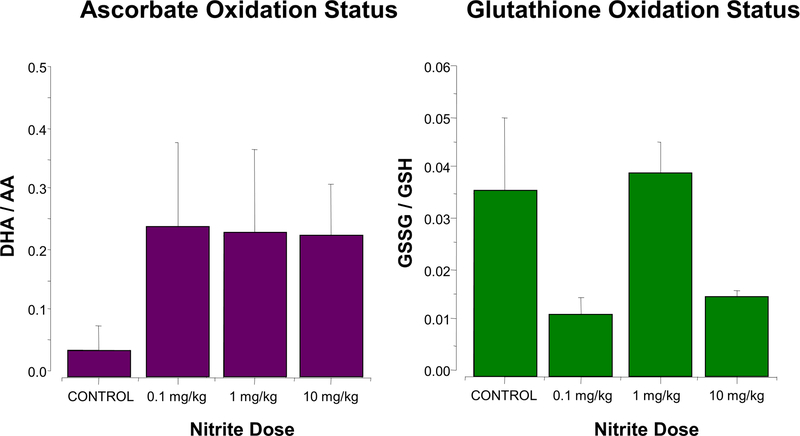
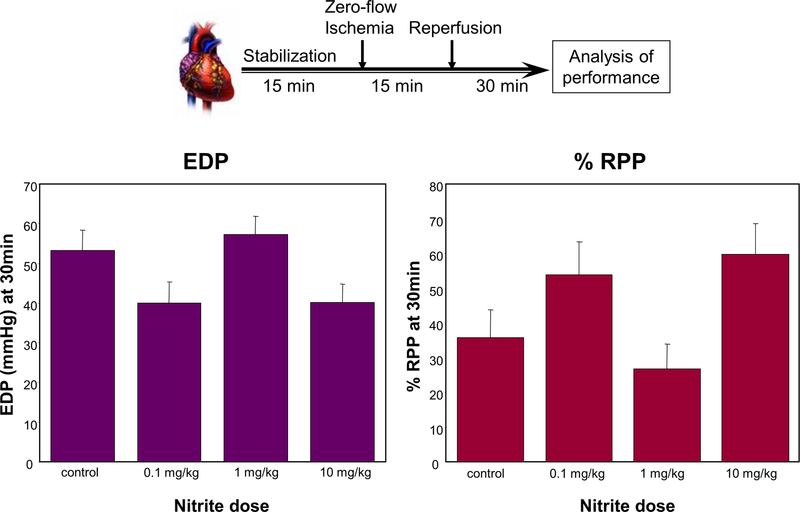
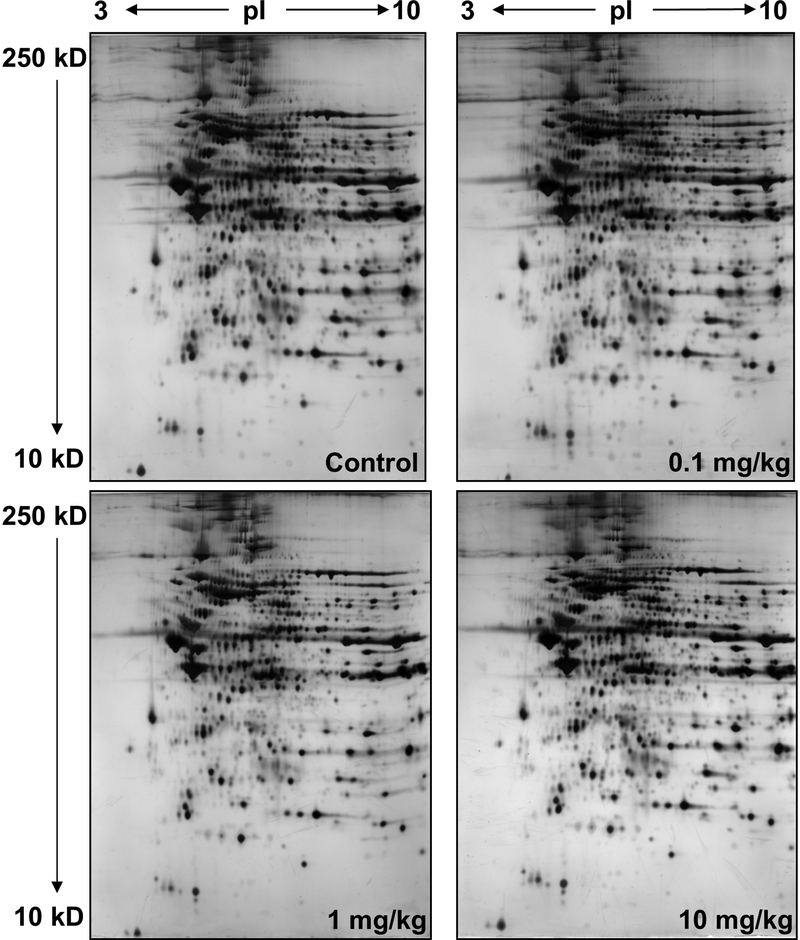
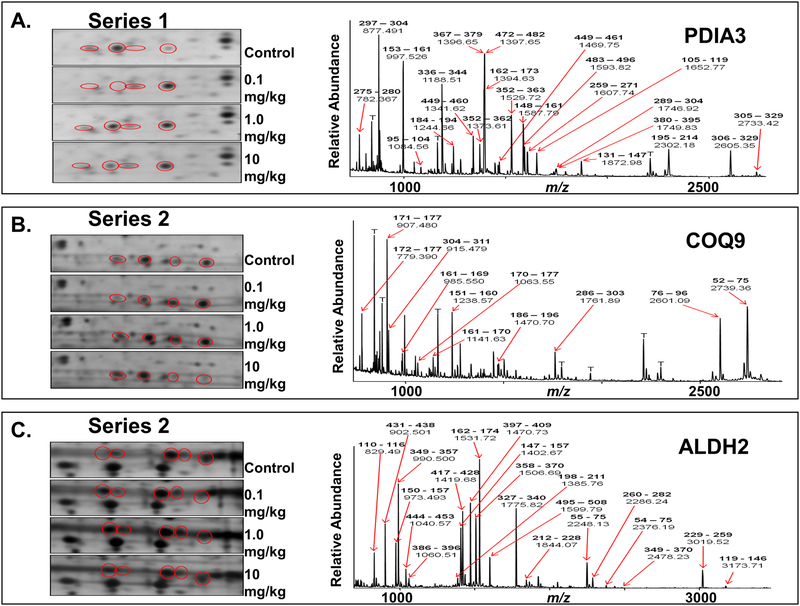
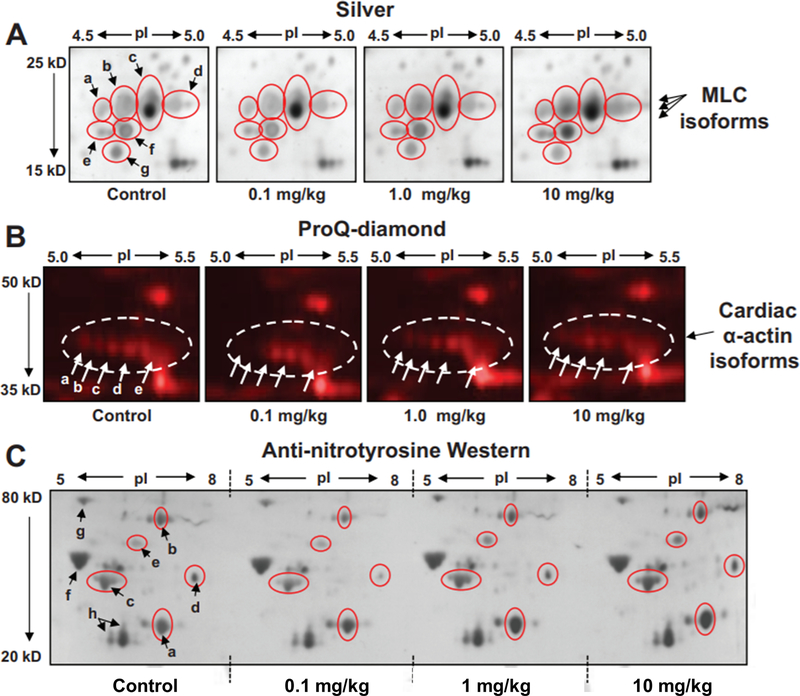
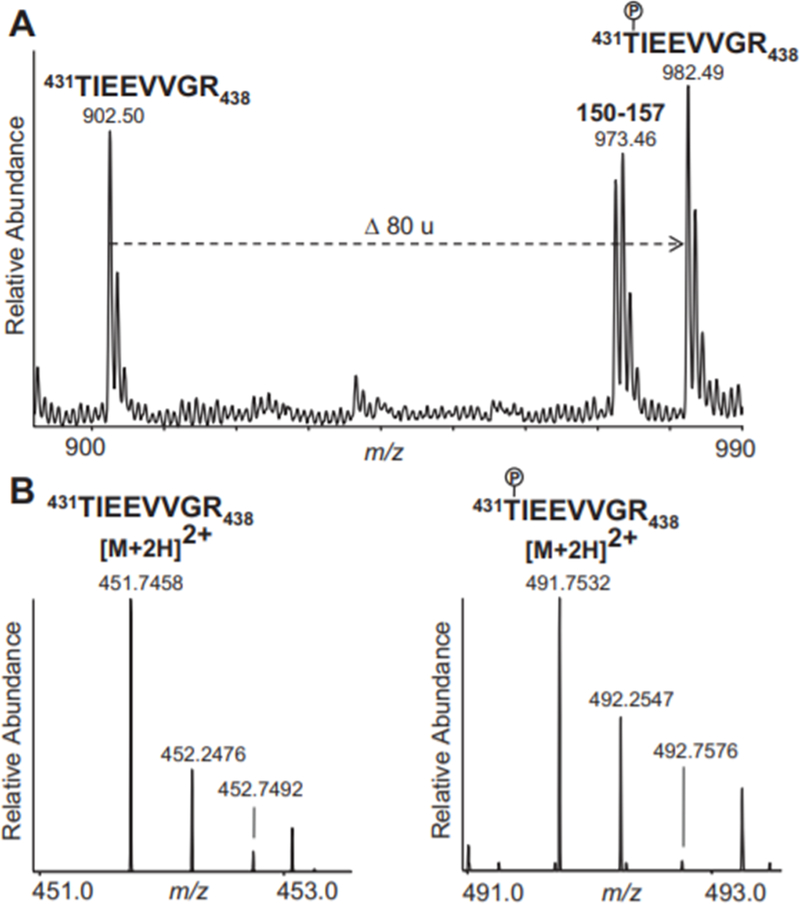
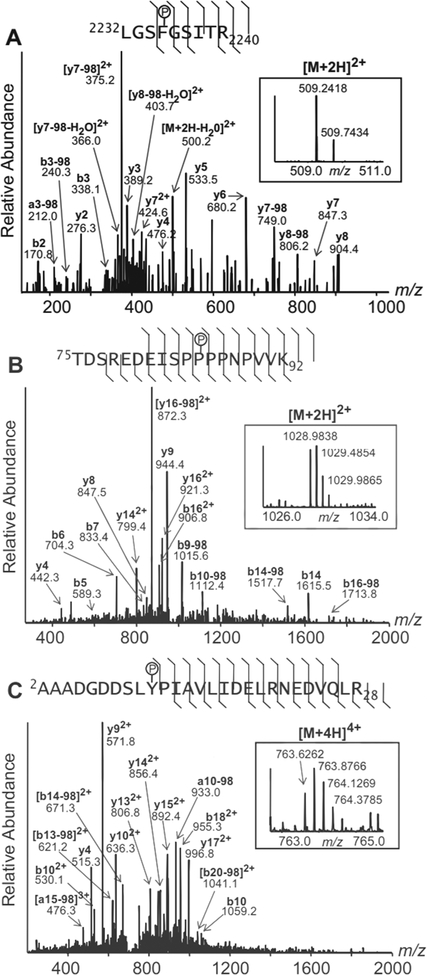
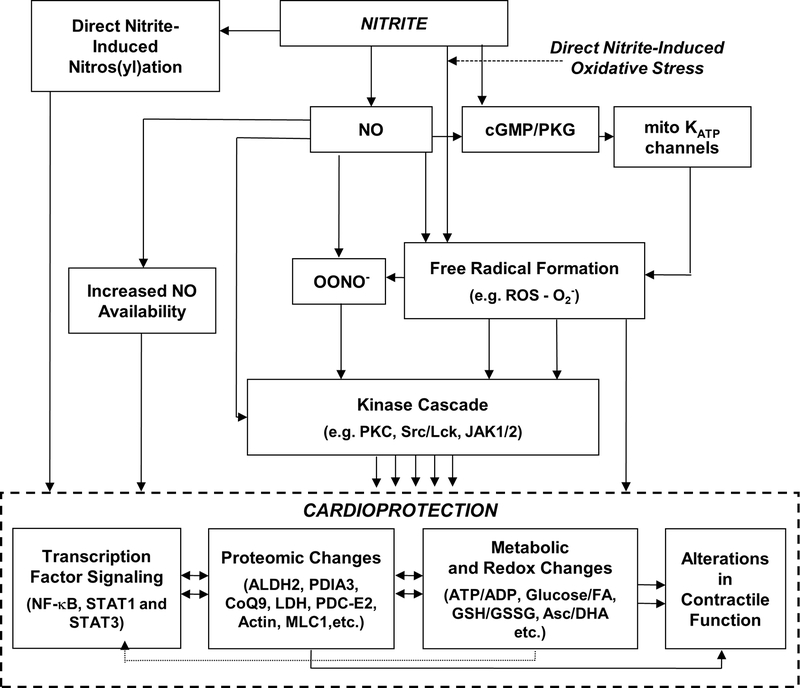
Nitrite has recently emerged as an important bioactive molecule, capable of conferring cardioprotection and a variety of other benefits in the cardiovascular system and elsewhere. The mechanisms by which it accomplishes these functions remain largely unclear. To characterize the dose response and corresponding cardiac sequelae of transient systemic elevations of nitrite, we assessed the time course of oxidation/nitros(yl)ation, as well as the metabolomic, proteomic, and associated functional changes in rat hearts following acute exposure to nitrite in vivo. Transient systemic nitrite elevations resulted in: (1) rapid formation of nitroso and nitrosyl species; (2) moderate short-term changes in cardiac redox status; (3) a pronounced increase in selective manifestations of long-term oxidative stress as evidenced by cardiac ascorbate oxidation, persisting long after changes in nitrite-related metabolites had normalized; (4) lasting reductions in glutathione oxidation (GSSG/GSH) and remarkably concordant nitrite-induced cardioprotection, which both followed a complex dose-response profile; and (5) significant nitrite-induced protein modifications (including phosphorylation) revealed by mass spectrometry-based proteomic studies. Altered proteins included those involved in metabolism (eg, aldehyde dehydrogenase 2, ubiquinone biosynthesis protein CoQ9, lactate dehydrogenase B), redox regulation (eg, protein disulfide isomerase A3), contractile function (eg, filamin-C), and serine/threonine kinase signaling (eg, protein kinase A R1alpha, protein phosphatase 2A A R1-alpha). Thus, brief elevations in plasma nitrite trigger a concerted cardioprotective response characterized by persistent changes in cardiac metabolism, redox stress, and alterations in myocardial signaling. These findings help elucidate possible mechanisms of nitrite-induced cardioprotection and have implications for nitrite dosing in therapeutic regimens.
Figures
References
-
- Mazzone M, Carmeliet P. Drug discovery: a lifeline for suffocating tissues. Nature. 2008;453:1194–1195. - PubMed
-
- Zweier JL, Wang P, Samouilov A, Kuppusamy P. Enzyme-independent formation of nitric oxide in biological tissues. Nat Med. 1995;1:804–809. - PubMed
-
- Lundberg JO, Weitzberg E, Gladwin MT. The nitrate-nitrite-nitric oxide pathway in physiology and therapeutics. Nat Rev Drug Discov. 2008;7:156–167. - PubMed
-
- Cosby K, Partovi KS, Crawford JH, Patel RP, Reiter CD, Martyr S, Yang BK, Waclawiw MA, Zalos G, Xu X, Huang KT, Shields H, Kim-Shapiro DB, Schechter AN, Cannon RO III, Gladwin MT. Nitrite reduction to nitric oxide by deoxyhemoglobin vasodilates the human circulation. Nat Med. 2003;9:1498–1505. - PubMed
-
- Maher AR, Milsom AB, Gunaruwan P, Abozguia K, Ahmed I, Weaver RA, Thomas P, Ashrafian H, Born GV, James PE, Frenneaux MP. Hypoxic modulation of exogenous nitrite-induced vasodilation in humans. Circulation. 2008;117:670–677. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 HL069029/HL/NHLBI NIH HHS/United States
- S10 RR20946/RR/NCRR NIH HHS/United States
- P41 RR10888/RR/NCRR NIH HHS/United States
- S10 RR020946/RR/NCRR NIH HHS/United States
- R01 HL69029/HL/NHLBI NIH HHS/United States
- N01HV28178/HV/NHLBI NIH HHS/United States
- R21 DA020644/DA/NIDA NIH HHS/United States
- S10 RR15942/RR/NCRR NIH HHS/United States
- P41 RR010888/RR/NCRR NIH HHS/United States
- P41 GM104603/GM/NIGMS NIH HHS/United States
- N01 HV028178/HV/NHLBI NIH HHS/United States
- G0701115/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
