

Identification of potential pathway mediation targets in Toll-like receptor signaling
- PMID: 19229310
- PMCID: PMC2634968
- DOI: 10.1371/journal.pcbi.1000292
Identification of potential pathway mediation targets in Toll-like receptor signaling
Erratum in
- PLoS Comput Biol. 2009 Nov;5(11). doi: 10.1371/annotation/5cc0d918-83b8-44e4-9778-b96a249d4099
Abstract
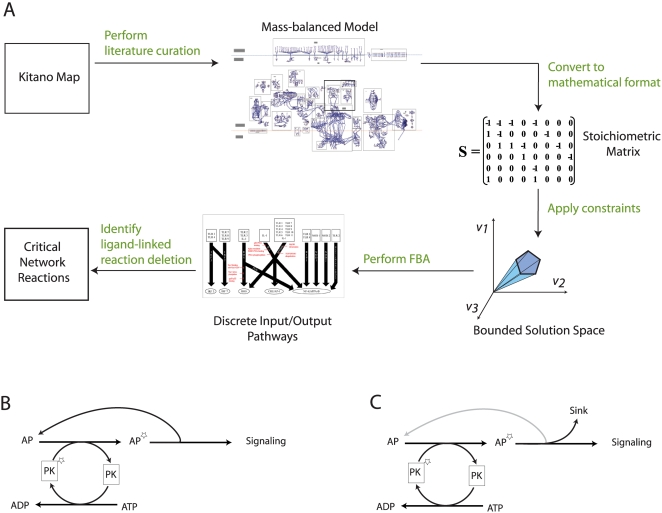
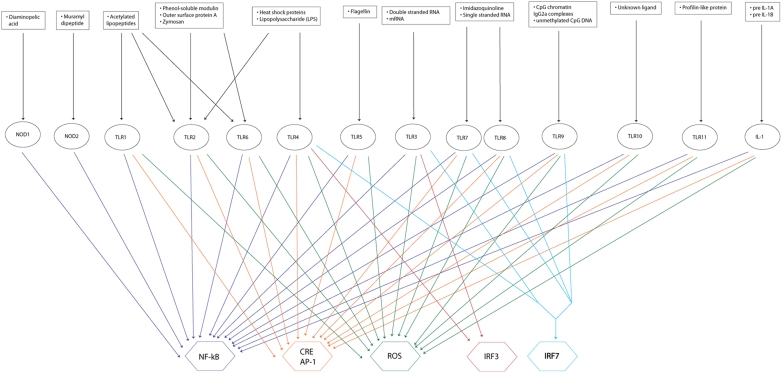
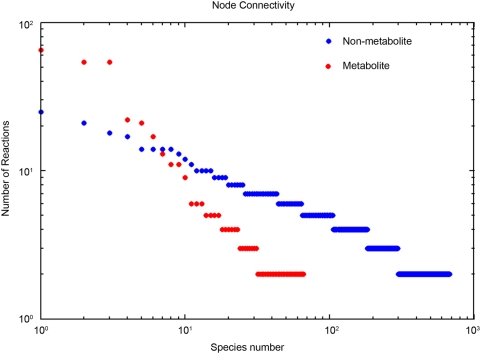
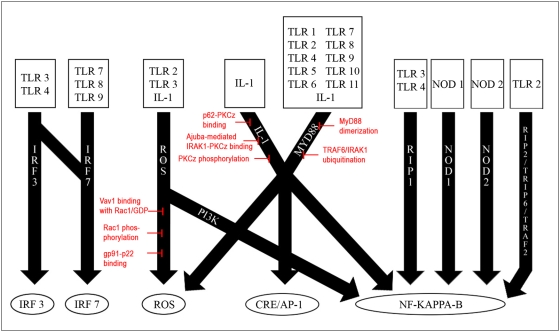
Recent advances in reconstruction and analytical methods for signaling networks have spurred the development of large-scale models that incorporate fully functional and biologically relevant features. An extended reconstruction of the human Toll-like receptor signaling network is presented herein. This reconstruction contains an extensive complement of kinases, phosphatases, and other associated proteins that mediate the signaling cascade along with a delineation of their associated chemical reactions. A computational framework based on the methods of large-scale convex analysis was developed and applied to this network to characterize input-output relationships. The input-output relationships enabled significant modularization of the network into ten pathways. The analysis identified potential candidates for inhibitory mediation of TLR signaling with respect to their specificity and potency. Subsequently, we were able to identify eight novel inhibition targets through constraint-based modeling methods. The results of this study are expected to yield meaningful avenues for further research in the task of mediating the Toll-like receptor signaling network and its effects.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
Similar articles
-
Signaling to NF-kappaB by Toll-like receptors.Trends Mol Med. 2007 Nov;13(11):460-9. doi: 10.1016/j.molmed.2007.09.002. Epub 2007 Oct 29. Trends Mol Med. 2007. PMID: 18029230 Review.
-
Ubiquitination and de-ubiquitination: role in regulation of signaling by Toll-like receptors.J Endotoxin Res. 2006;12(6):337-45. doi: 10.1179/096805106X118915. J Endotoxin Res. 2006. PMID: 17254387 Review.
-
Combined CpG and poly I:C stimulation of monocytes results in unique signaling activation not observed with the individual ligands.Cell Signal. 2013 Nov;25(11):2246-54. doi: 10.1016/j.cellsig.2013.07.014. Epub 2013 Jul 19. Cell Signal. 2013. PMID: 23876795
-
Peptides targeting Toll-like receptor signalling pathways for novel immune therapeutics.Curr Pharm Des. 2010;16(9):1063-80. doi: 10.2174/138161210790963841. Curr Pharm Des. 2010. PMID: 20030612 Review.
-
Therapeutic targeting of pattern-recognition receptors.Int Immunopharmacol. 2006 Jun;6(6):863-9. doi: 10.1016/j.intimp.2006.02.005. Epub 2006 Mar 3. Int Immunopharmacol. 2006. PMID: 16644471 Review.
Cited by
-
A protocol for generating a high-quality genome-scale metabolic reconstruction.Nat Protoc. 2010 Jan;5(1):93-121. doi: 10.1038/nprot.2009.203. Epub 2010 Jan 7. Nat Protoc. 2010. PMID: 20057383 Free PMC article.
-
Towards a Systems Biology Approach to Understanding the Lichen Symbiosis: Opportunities and Challenges of Implementing Network Modelling.Front Microbiol. 2021 May 3;12:667864. doi: 10.3389/fmicb.2021.667864. eCollection 2021. Front Microbiol. 2021. PMID: 34012428 Free PMC article. Review.
-
A comprehensive map of the influenza A virus replication cycle.BMC Syst Biol. 2013 Oct 2;7:97. doi: 10.1186/1752-0509-7-97. BMC Syst Biol. 2013. PMID: 24088197 Free PMC article.
-
Quantitative prediction of cellular metabolism with constraint-based models: the COBRA Toolbox v2.0.Nat Protoc. 2011 Aug 4;6(9):1290-307. doi: 10.1038/nprot.2011.308. Nat Protoc. 2011. PMID: 21886097 Free PMC article.
-
Information processing by biochemical networks: a dynamic approach.J R Soc Interface. 2011 Feb 6;8(55):186-200. doi: 10.1098/rsif.2010.0287. Epub 2010 Aug 4. J R Soc Interface. 2011. PMID: 20685691 Free PMC article.
References
-
- Kaisho T, Akira S. Toll-like receptor function and signaling. J Allergy Clin Immunol. 2006;117:979–987; quiz 988. - PubMed
-
- Hoebe K, Jiang Z, Georgel P, Tabeta K, Janssen E, et al. TLR signaling pathways: opportunities for activation and blockade in pursuit of therapy. Curr Pharm Des. 2006;12:4123–4134. - PubMed
-
- Trinchieri G, Sher A. Cooperation of Toll-like receptor signals in innate immune defence. Nat Rev Immunol. 2007;7:179–190. - PubMed
-
- Mullick AE, Tobias PS, Curtiss LK. Toll-like receptors and atherosclerosis: key contributors in disease and health? Immunol Res. 2006;34:193–209. - PubMed
-
- Stoll LL, Denning GM, Weintraub NL. Endotoxin, TLR4 signaling and vascular inflammation: potential therapeutic targets in cardiovascular disease. Curr Pharm Des. 2006;12:4229–4245. - PubMed
