

Characterization and coding of behaviorally significant odor mixtures
- PMID: 19230669
- PMCID: PMC2677194
- DOI: 10.1016/j.cub.2009.01.041
Characterization and coding of behaviorally significant odor mixtures
Abstract
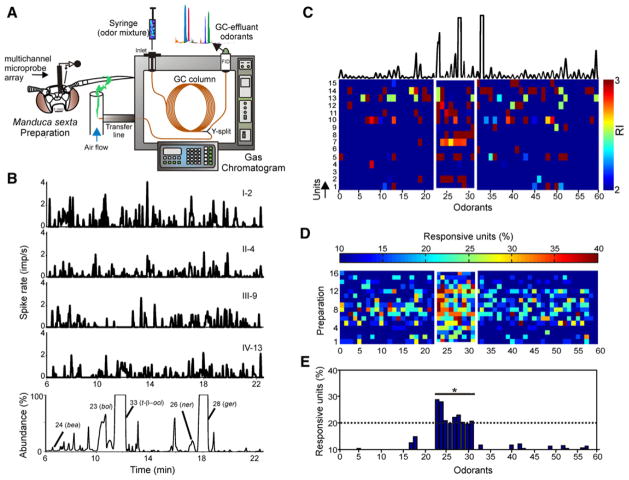
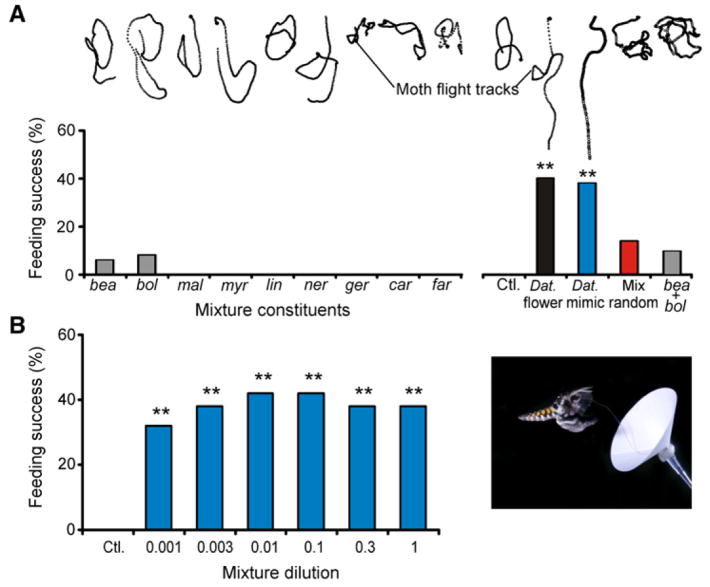
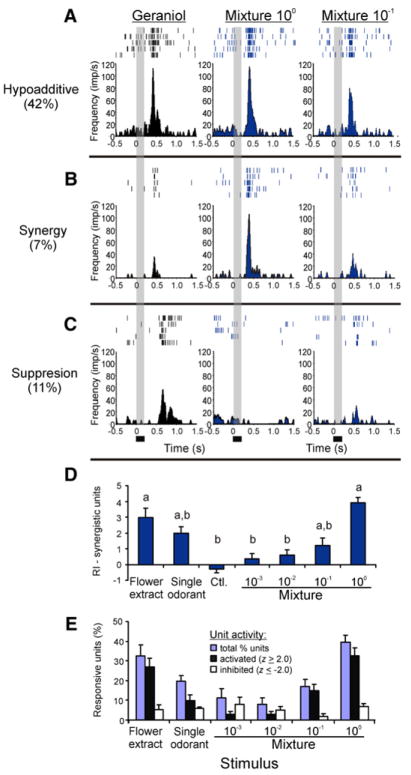
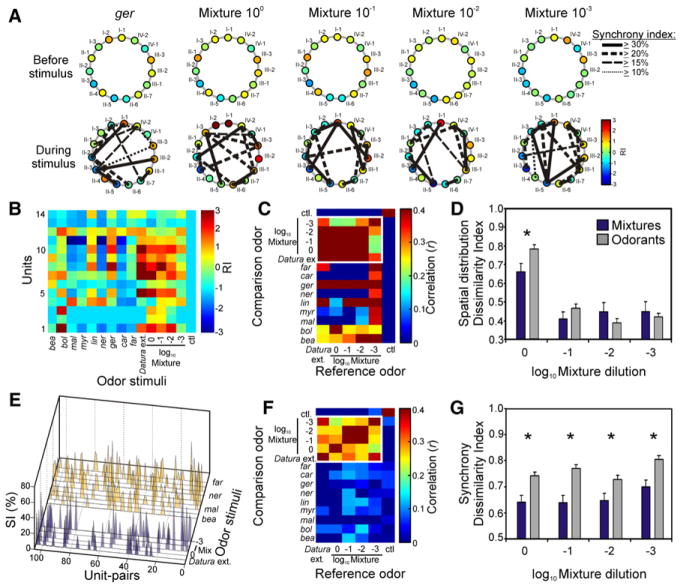
For animals to execute odor-driven behaviors, the olfactory system must process complex odor signals and maintain stimulus identity in the face of constantly changing odor intensities [1-5]. Surprisingly, how the olfactory system maintains identity of complex odors is unclear [6-10]. We took advantage of the plant-pollinator relationship between the Sacred Datura (Datura wrightii) and the moth Manduca sexta[11, 12] to determine how olfactory networks in this insect's brain represent odor mixtures. We combined gas chromatography and neural-ensemble recording in the moth's antennal lobe to examine population codes for the floral mixture and its fractionated components. Although the floral scent of D. wrightii comprises at least 60 compounds, only nine of those elicited robust neural responses. Behavioral experiments confirmed that these nine odorants mediate flower-foraging behaviors, but only as a mixture. Moreover, the mixture evoked equivalent foraging behaviors over a 1000-fold range in dilution, suggesting a singular percept across this concentration range. Furthermore, neural-ensemble recordings in the moth's antennal lobe revealed that reliable encoding of the floral mixture is organized through synchronized activity distributed across a population of glomerular coding units, and this timing mechanism may bind the features of a complex stimulus into a coherent odor percept.
Figures
References
-
- Bartelt RJ, Schaner AM, Jackson LL. cis-Vaccenyl acetate as an aggregation pheromone in Drosophila melanogaster. J Chem Ecol. 1985;11:1747–1756. - PubMed
-
- Boch R, Shearer DA. Identification of geraniol as the active component in the Nassanoff pheromone of the honey bee. Nature. 1962;194:704–706. - PubMed
-
- Kaissling KE. Peripheral mechanisms of pheromone reception in moths. Chem Senses. 1996;21:257–268. - PubMed
-
- Laska M, Fendt M, Wieser A, Endres T, Hernandez Salazar LT, Apfelbach R. Detecting danger – or just another odorant? Olfactory sensitivity for the fox odor component 2,4,5-trimethylthiazoline in four species of mammals. Physiol Behav. 2005;84:211–215. - PubMed
-
- Williams IH, Pickett JA, Martin AP. The Nasonov pheromone of the honey bee Apis mellifera L. (Hymenoptera, Apidae). Part II Bioassay of the components using foragers. J Chem Ecol. 1981;7:225–237. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
