

Review
doi: 10.1146/annurev.biochem.78.082907.145923.
Multidrug resistance in bacteria
Affiliations
- PMID: 19231985
- PMCID: PMC2839888
- DOI: 10.1146/annurev.biochem.78.082907.145923
Item in Clipboard
Review
Multidrug resistance in bacteria
Annu Rev Biochem.
2009.
Abstract
Large amounts of antibiotics used for human therapy, as well as for farm animals and even for fish in aquaculture, resulted in the selection of pathogenic bacteria resistant to multiple drugs. Multidrug resistance in bacteria may be generated by one of two mechanisms. First, these bacteria may accumulate multiple genes, each coding for resistance to a single drug, within a single cell. This accumulation occurs typically on resistance (R) plasmids. Second, multidrug resistance may also occur by the increased expression of genes that code for multidrug efflux pumps, extruding a wide range of drugs. This review discusses our current knowledge on the molecular mechanisms involved in both types of resistance.
Figures
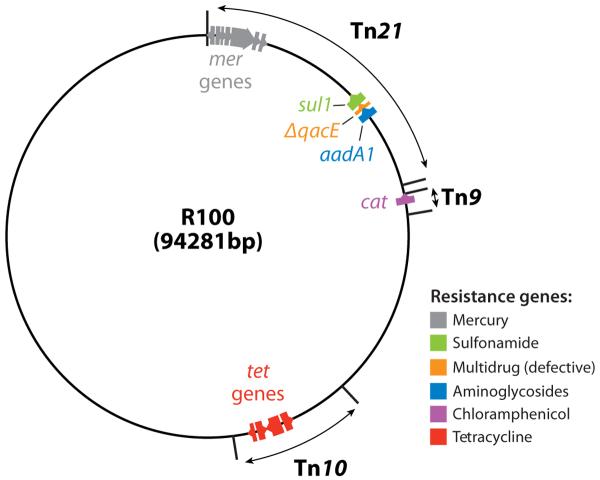
Map of an early R plasmid R100. Tetracycline resistance gene tetA is in the transposon Tn10, chloramphenicol acetyltransferase (cat) as a part of Tn9, and sulfonamide resistance gene sul1 and an aminoglycoside adenyltransferase gene aadA1 as a part of the large transposon Tn21. The resistance genes are color coded, and the direction of transcription is shown by the arrows. Much of the blank areas outside the transposons are occupied by the transfer genes, needed for the formation of relaxosomes, the coupling factor, and the mating-pair formation. This figure is based on the nucleotide sequence deposited by (30) and GenBank sequence NC 002134, submitted by G. Sempei and K. Mizobuchi. ΔqacE is a defective version of a gene coding for a multidrug efflux pump of Smr family (see text).
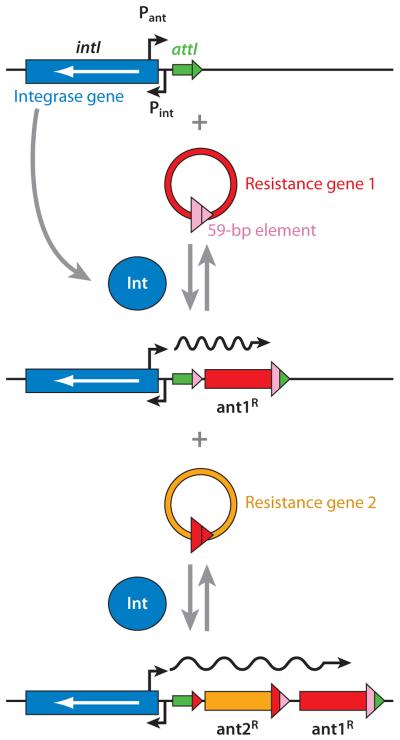
Presumed mechanism of resistance gene capture by integrons. An integron contains the gene for site-specific integrase (blue) and the specific integration site attI (green). When the resistance gene 1 (red) in the circular cassette form containing the 3′-terminal 59-bp element (pink) is available, the gene is integrated at the attI site, regenerating a slightly altered attI sequence (now in green and pink). This can then accept the second resistance gene casette, and the process can go on in this manner. The resistance genes, all in the same orientation, are transcribed by the powerful promoter provided by the integron (Pant). From Reference 159 with permission from Elsevier Ltd.
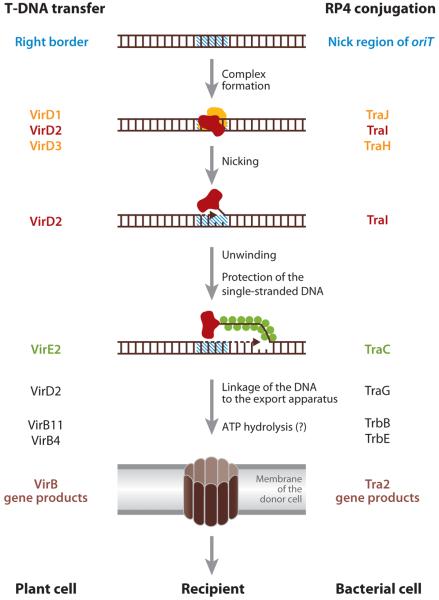
Steps in T-DNA transfer in Agrobacterium tumefaciens (annotation on the left) and conjugative plasmid DNA transfer in enteric bacteria (annotation on the right). From Reference 41 with permission from Elsevier Ltd.
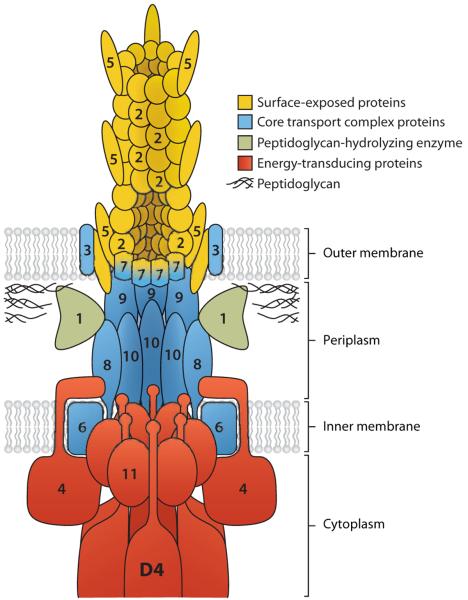
A model of the mating-pair formation complex with the coupling protein (VirD4). The proteins are named after the VirB components in Agrobacterium tumefaciens, but homologs for most of these proteins are found in the conjugative R plasmid transfer systems. Number n denotes the product of virBn gene. The virD4 product is shown as D4. From Reference 39 with permission from Elsevier Ltd.
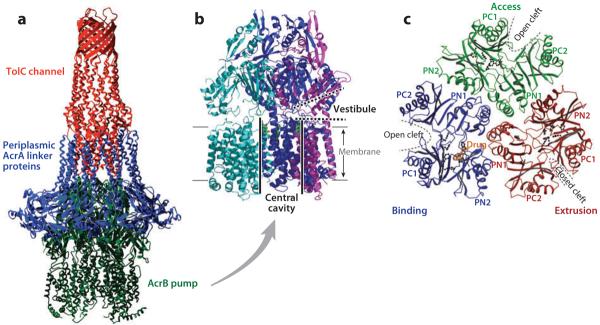
(a) The model of the AcrB-AcrA-TolC tripartite complex that exports drugs directly into the medium. The transmembrane domain of the AcrB pump trimer is embedded in the cytoplasmic membrane, whereas its periplasmic domain is connected to the TolC channel through a number of periplasmic AcrA linker proteins. From Reference 105 with permission from Elsevier Ltd. (b) AcrB trimer. Each protomer is shown in a different color. The large central cavity (thick black lines) is connected to the periplasm through vestibules (thick dotted lines) between protomers. The proximal portion of the structure was cut away to reveal the presence of vestibule. Drawn by using PyMol with Protein Data Bank coordinate file 1OYE. (c) The periplasmic domain of the asymmetric AcrB trimer viewed from the top. The conformation of each protomer is characteristic, with open or closed external clefts. A drug molecule is seen bound to the binding protomer. From Reference 113 with permission from Nature Publishing Group.
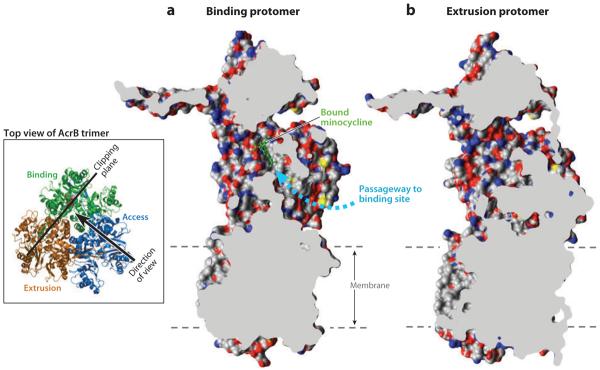
Cut out view of the binding protomer with the bound minocycline (in a ball-and-stick representation in green) (a) and the extruding protomer (b), both from Protein Data Bank file 2DRD. Molecular graphics images were produced using the UCSF Chimera package from the Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco (supported by NIH P41 RR-01081) (119). The wide passageway from the external surface to the binding site (dashed arrow) seen in (a) appears to be completely closed in (b). The dashed lines show the approximate limits of the membrane bilayer. Modified after (120) with permission from Elsevier. The inset shows the location and direction of the clipping plane as well as the direction of the view in (a). The binding, extrusion, and access protomers are indicated.
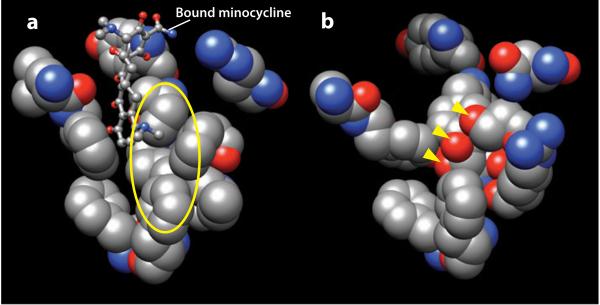
Substrate binding pockets of AcrB and a homology-modeled AcrD. (a) Residues facing the bound minocycline (in a ball-and-stick model), i.e., residues 177, 178, 275–278, 610, 612, 615, 620, 626, and 628 of the binding protomer of AcrB in Protein Data Bank file 2DRD are shown in space-filling models. The walls are remarkably hydrophobic, as seen for example in the area covered by the yellow ellipse. (b) Hypothetical structure of the corresponding region in AcrD. AcrD structure was built by homology modeling, using the binding protomer of AcrB as the template. The area covered by the ellipse in (a) is now seen to be studded with oxygen atoms. The oxygen atoms shown by the arrowheads belong to the side chains of, from top, Ser replacing Phe617 of AcrB, Tyr replacing Ile277, and Thr replacing Phe611. Molecular graphics images were produced using the UCSF Chimera package from the Resource for Biocomputing, Visualization, and Informatics at the University of California, San Francisco (supported by NIH P41 RR-01081) (119).
References
-
- Livermore DM. The need for new antibiotics. Clin. Microbiol. Infect. 2004;10(Suppl 4):1–9. - PubMed
-
- Hooper DC. Mechanisms of action and resistance of older and newer fluoroquinolones. Clin. Infect. Dis. 2000;31(Suppl 2):S24–28. - PubMed
-
- Poehlsgaard J, Douthwaite S. The bacterial ribosome as a target for antibiotics. Nat. Rev. Microbiol. 2005;3:870–81. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
