

Contrast enhancement of stimulus intermittency in a primary olfactory network and its behavioral significance
- PMID: 19232128
- PMCID: PMC2687775
- DOI: 10.1186/jbiol120
Contrast enhancement of stimulus intermittency in a primary olfactory network and its behavioral significance
Abstract
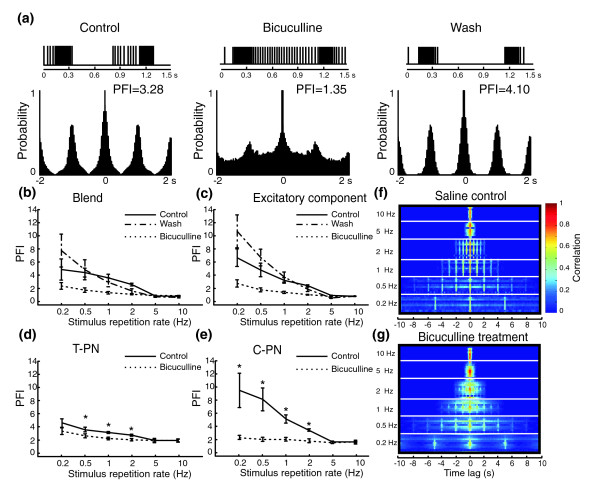
Background: An animal navigating to an unseen odor source must accurately resolve the spatiotemporal distribution of that stimulus in order to express appropriate upwind flight behavior. Intermittency of natural odor plumes, caused by air turbulence, is critically important for many insects, including the hawkmoth, Manduca sexta, for odor-modulated search behavior to an odor source. When a moth's antennae receive intermittent odor stimulation, the projection neurons (PNs) in the primary olfactory centers (the antennal lobes), which are analogous to the olfactory bulbs of vertebrates, generate discrete bursts of action potentials separated by periods of inhibition, suggesting that the PNs may use the binary burst/non-burst neural patterns to resolve and enhance the intermittency of the stimulus encountered in the odor plume.
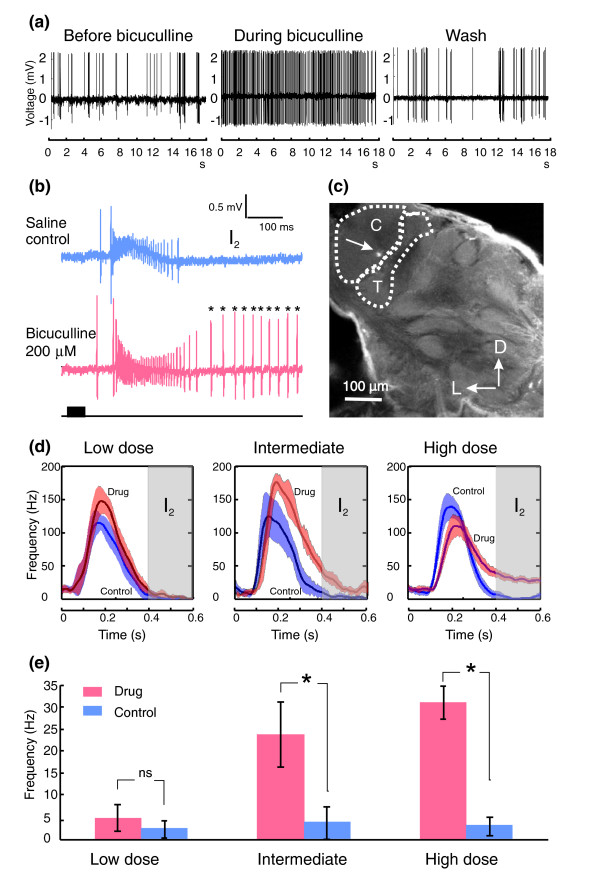
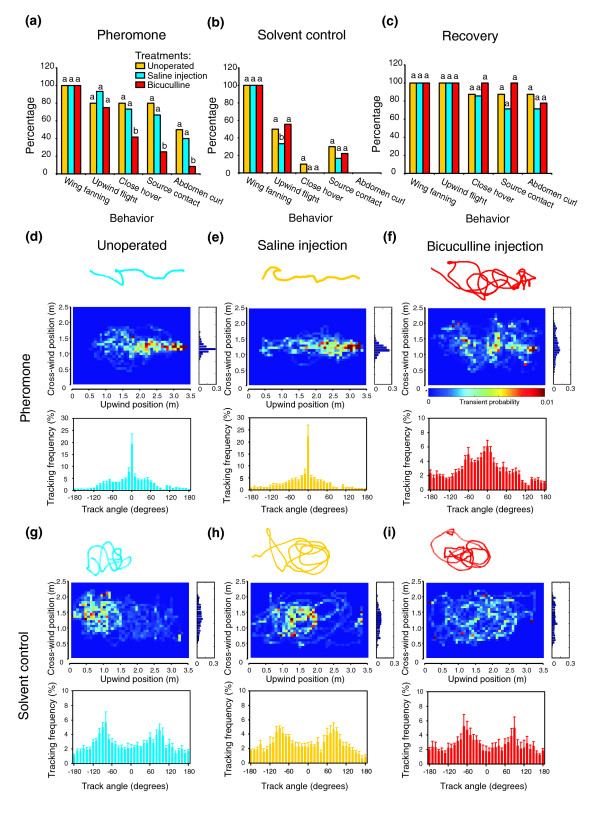
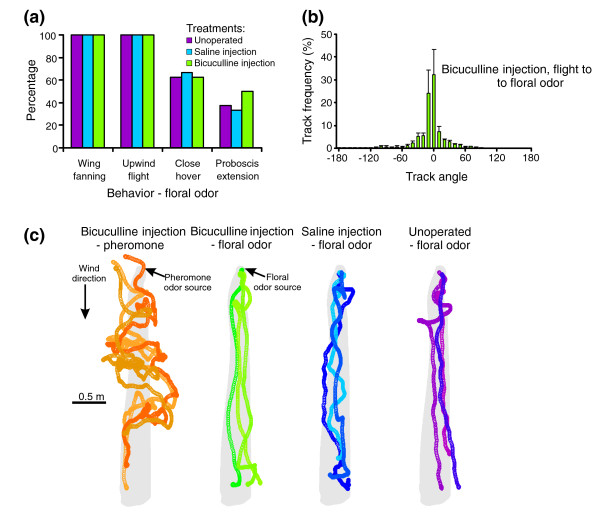
Results: We tested this hypothesis first by establishing that bicuculline methiodide reliably and reversibly disrupted the ability of PNs to produce bursting response patterns. Behavioral studies, in turn, demonstrated that after injecting this drug into the antennal lobe at the effective concentration used in the physiological experiments animals could no longer efficiently locate the odor source, even though they had detected the odor signal.
Conclusions: Our results establish a direct link between the bursting response pattern of PNs and the odor-tracking behavior of the moth, demonstrating the behavioral significance of resolving the dynamics of a natural odor stimulus in antennal lobe circuits.
Figures
Comment in
-
Representations of odor plume flux are accentuated deep within the moth brain.J Biol. 2009;8(2):16. doi: 10.1186/jbiol122. Epub 2009 Feb 20. J Biol. 2009. PMID: 19239722 Free PMC article. Review.
References
-
- Adrian ED. Electrical activity of the nervous system. Arch Neurol Psychiatry. 1934;32:1125–1136.
-
- Murlis J, Jones CD. Fine-scale structure of odor plumes in relation to insect orientation to distant pheromone and other attractant sources. Physiol Entomol. 1981;6:71–86. doi: 10.1111/j.1365-3032.1981.tb00262.x. - DOI
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
