

Review
doi: 10.1242/dev.020966.
Developmental roles of the histone lysine demethylases
Affiliations
- PMID: 19234061
- PMCID: PMC2692332
- DOI: 10.1242/dev.020966
Item in Clipboard
Review
Developmental roles of the histone lysine demethylases
Development.
2009 Mar.
Abstract
Since the discovery of the first histone lysine demethylase in 2004, two protein families with numerous members have been identified that demethylate various histone lysine residues. Initial studies of the histone lysine demethylases focused on their in vitro enzymatic activity but, more recently, model organisms have been used to examine the roles of these enzymes in vivo. Here, we review recent insights into the roles of the histone lysine demethylases in multiple aspects of development across various species, including in germline maintenance and meiosis, in early embryonic development and differentiation, and in hormone receptor-mediated transcriptional regulation.
Figures
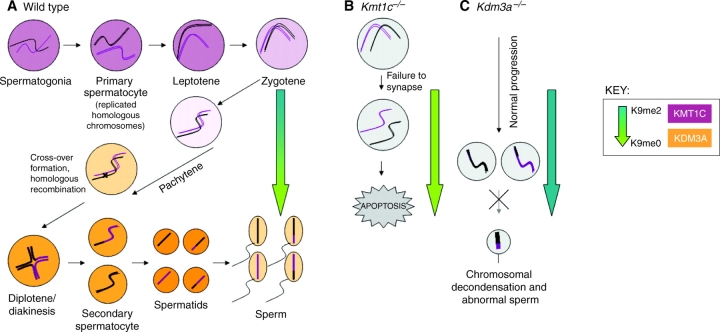
Histone H3 lysine 9 (H3K9) methylation balance in spermatogenesis. (A) H3K9me methyltransferases and demethylases are temporally regulated during spermatogenesis. In wild-type mice, mitotically cycling spermatogonia enter meiosis as primary spermatocytes, which then proceed through stages of early prophase I with concurrently high levels of the H3K9me histone lysine methyltransferase (KMT) G9a/KMT1C (purple shading in nuclei). During leptotene and zygotene, the replicated homologous chromosomes pair and DNA double-strand break (DSB) formation is initiated. During the early-to-late pachytene progression, G9a/KMT1C expression is lost and the H3K9me2 histone lysine demethylase (KDM) JHDM2A/KDM3A is expressed (orange shading in nuclei). The relative levels of H3K9me2 are represented by the blue-to-green arrow: in the earlier stages of meiosis there is increased H3K9me2 (blue), which is dramatically reduced during pachytene while H3K9me0 levels increase (green), reflecting the change from G9a/KMT1C to JHDM2A/KDM3A expression. (B) KMT1C is required for early stages of spermatogenesis. In Kmt1c–/– mutant mice, normal H3K9me2 levels are never achieved (green arrow), homologs are unable to pair/synapse, DSB repair is dysregulated, and cells undergo apoptosis (Tachibana et al., 2007). (C) KDM3A is required for late stages of spermatogenesis. In Kdm3a–/– mice, H3K9me2 levels stay high throughout spermatogenesis (blue arrow), leading to later defects in chromatin condensation as manifested in the few sperm that complete maturation (Okada et al., 2007). Both Kmt1c–/– and Kdm3a–/– mutants are male-sterile.
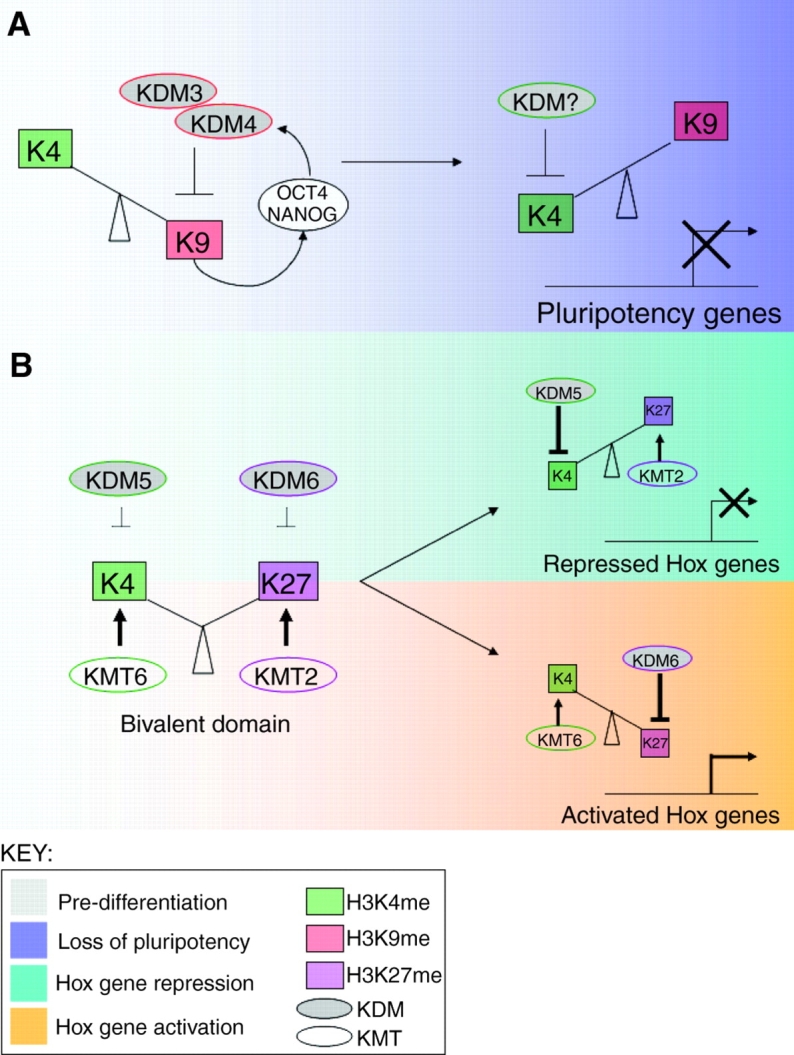
Opposing histone demethylases regulate developmental genes. (A) Model for demethylase regulation of pluripotency genes. Early in mouse development, the pluripotency-maintaining factors OCT4 and NANOG, and the H3K9me histone lysine demethylases KDM3 and KDM4, participate in a positive regulatory feedback loop. Upon differentiation, these demethylases are downregulated, allowing for increased H3K9 methylation and the repression of Oct4/Nanog. This model predicts that the activity of an H3K4me KDM is induced upon differentiation so as to remove the H3K4me modification from the pluripotency genes. LSD1/KDM1 is one candidate for this role, although this model needs to be experimentally confirmed. (B) Model for demethylase regulation of Hox genes. Early in mouse development, Hox genes are marked by a unique `bivalent domain', which features both H3K27me3 and H3K4me3. KDMs for both modifications are present but, for unknown reasons, do not demethylate these marks. Upon differentiation, these bivalent domains resolve into either H3K27me3 (repressed) or H3K4me3 (activated) via the actions of KDMs and KMTs.
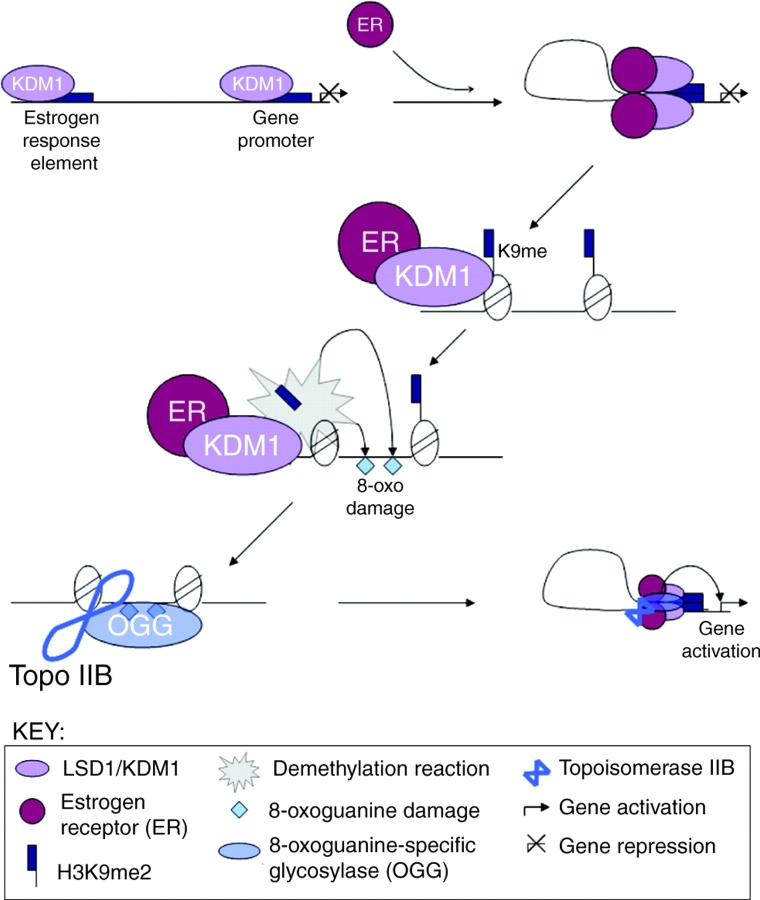
Model for LSD1/KDM1 regulation of estrogen receptor-induced gene expression. LSD1/KDM1 is constitutively associated with upstream estrogen-response elements (EREs) and promoters of estrogen receptor (ER) target genes. Upon addition of ligand (in the form of the hormone estrogen), the activated ER binds to both the enhancer and promoter, and triggers LSD1/KDM1-dependent DNA looping. LSD1/KDM1 then demethylates H3K9me2/1, and hydrogen peroxide generated by the demethylase reaction triggers localized 8-oxoguanine DNA damage. This damage recruits 8-oxoguanine-specific glycosylase (OGG) and topoisomerase IIβ (Topo IIβ), which in turn triggers chromatin conformational changes that promote gene transcription (Perillo et al., 2008).
References
-
- Agger, K., Cloos, P. A., Christensen, J., Pasini, D., Rose, S., Rappsilber, J., Issaeva, I., Canaani, E., Salcini, A. E. and Helin, K. (2007). UTX and JMJD3 are histone H3K27 demethylases involved in HOX gene regulation and development. Nature 449, 731-734. - PubMed
-
- Agger, K., Christensen, J., Cloos, P. A. and Helin, K. (2008). The emerging functions of histone demethylases. Curr. Opin. Genet. Dev. 18, 159-168. - PubMed
-
- Akam, M. (1995). Hox genes and the evolution of diverse body plans. Philos. Trans. R. Soc. Lond. B Biol. Sci. 349, 313-319. - PubMed
-
- Akimoto, C., Kitagawa, H., Matsumoto, T. and Kato, S. (2008). Spermatogenesis-specific association of SMCY and MSH5. Genes Cells 13, 623-633. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
