

Drought stress-induced Rma1H1, a RING membrane-anchor E3 ubiquitin ligase homolog, regulates aquaporin levels via ubiquitination in transgenic Arabidopsis plants
- PMID: 19234086
- PMCID: PMC2660634
- DOI: 10.1105/tpc.108.061994
Drought stress-induced Rma1H1, a RING membrane-anchor E3 ubiquitin ligase homolog, regulates aquaporin levels via ubiquitination in transgenic Arabidopsis plants
Abstract
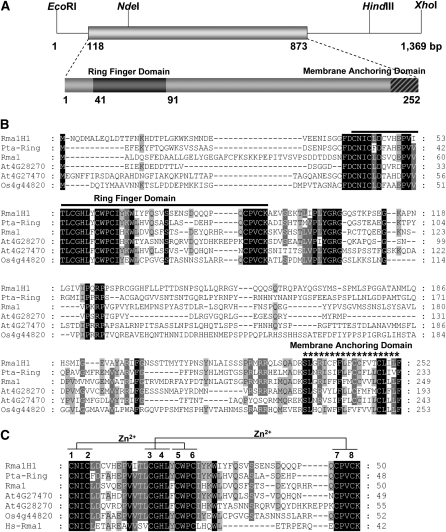
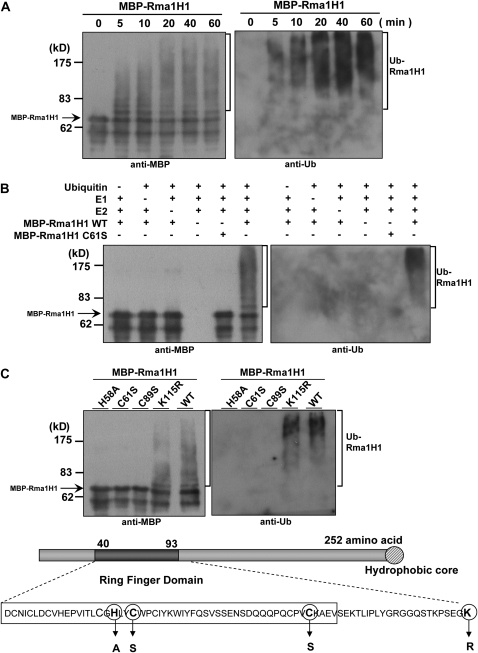
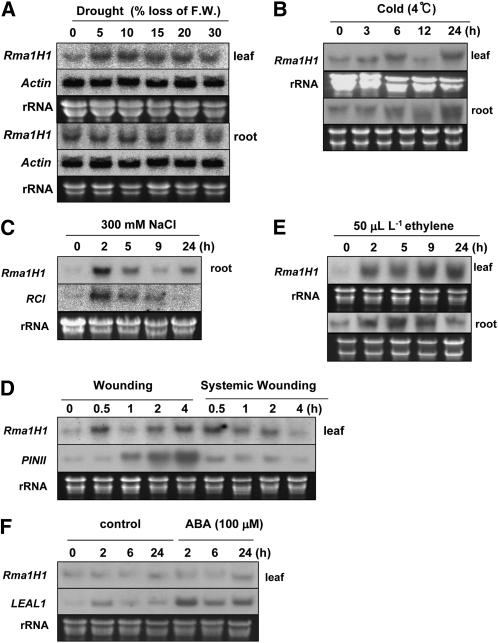
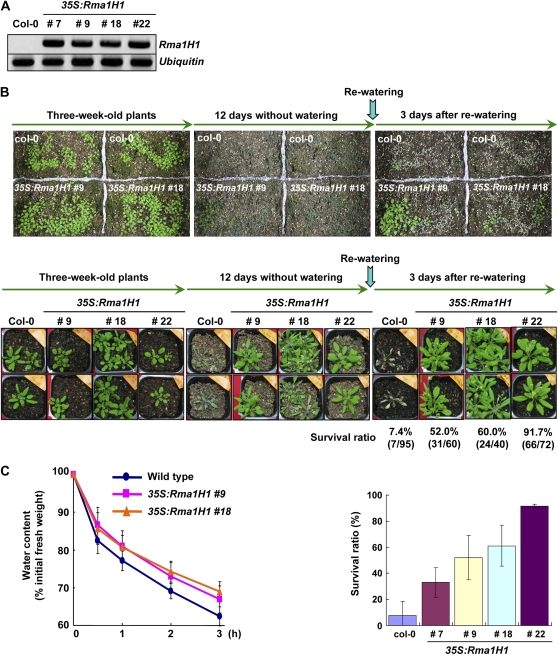
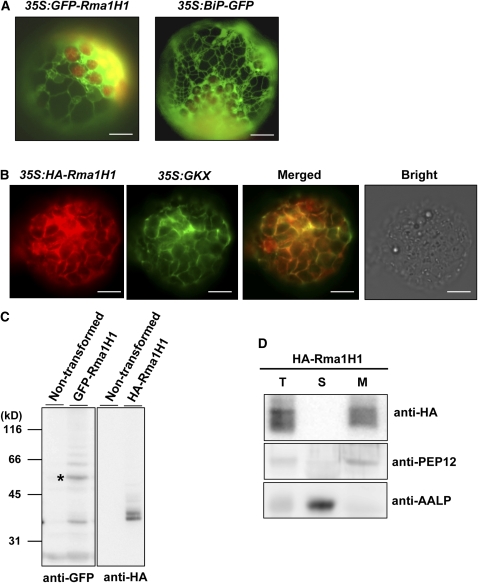
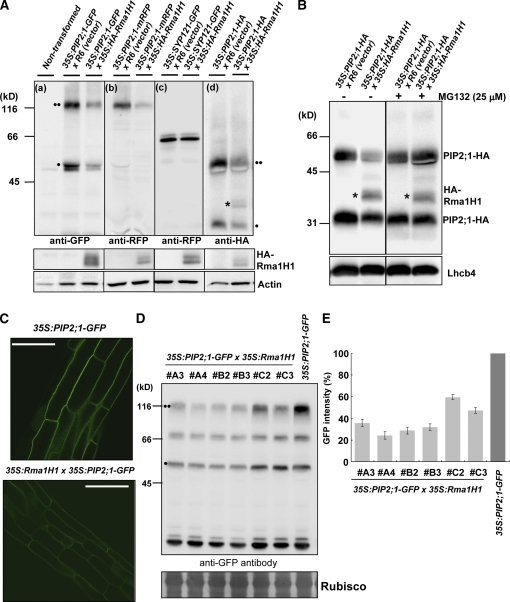
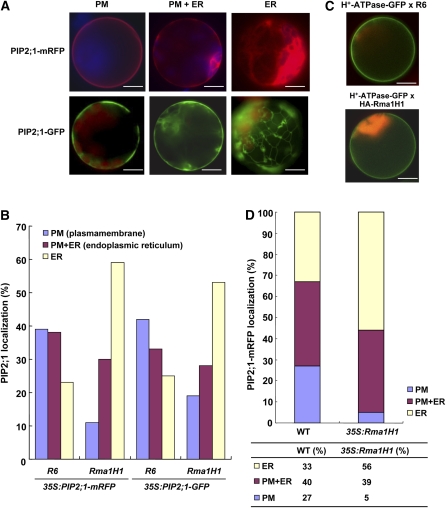
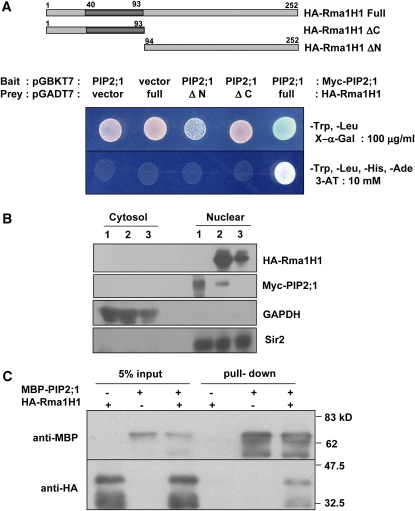
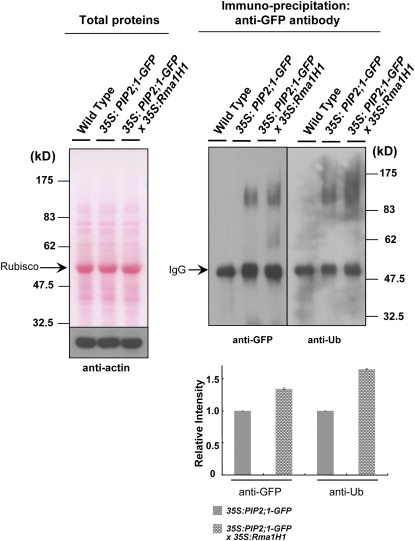
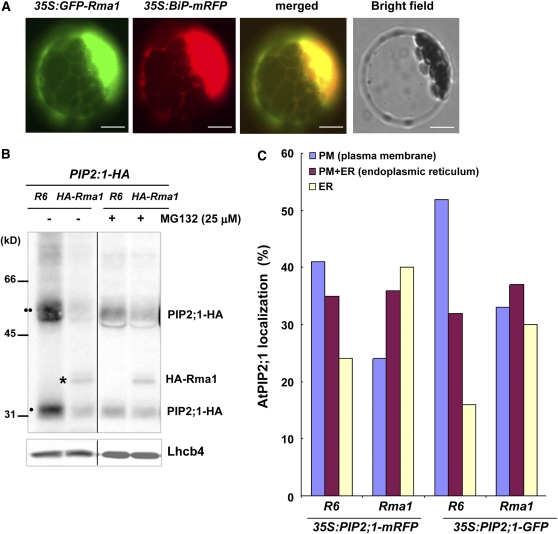
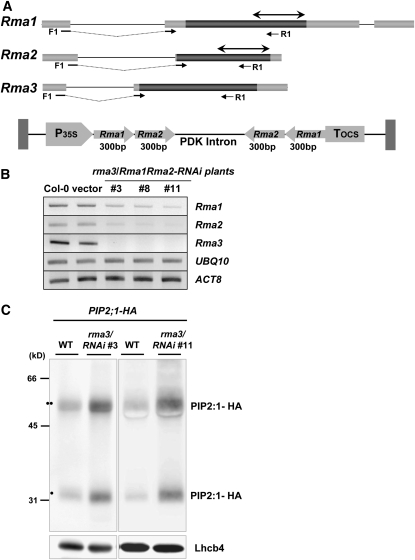
Ubiquitination is involved in a variety of biological processes, but the exact role of ubiquitination in abiotic responses is not clearly understood in higher plants. Here, we investigated Rma1H1, a hot pepper (Capsicum annuum) homolog of a human RING membrane-anchor 1 E3 ubiquitin (Ub) ligase. Bacterially expressed Rma1H1 displayed E3 Ub ligase activity in vitro. Rma1H1 was rapidly induced by various abiotic stresses, including dehydration, and its overexpression in transgenic Arabidopsis thaliana plants conferred strongly enhanced tolerance to drought stress. Colocalization experiments with marker proteins revealed that Rma1H1 resides in the endoplasmic reticulum (ER) membrane. Overexpression of Rma1H1 in Arabidopsis inhibited trafficking of an aquaporin isoform PIP2;1 from the ER to the plasma membrane and reduced PIP2;1 levels in protoplasts and transgenic plants. This Rma1H1-induced reduction of PIP2;1 was inhibited by MG132, an inhibitor of the 26S proteasome. Furthermore, Rma1H1 interacted with PIP2;1 in vitro and ubiquitinated it in vivo. Similar to Rma1H1, Rma1, an Arabidopsis homolog of Rma1H1, localized to the ER, and its overexpression reduced the PIP2;1 protein level and inhibited trafficking of PIP2;1 from the ER to the plasma membrane in protoplasts. In addition, reduced expression of Rma homologs resulted in the increased level of PIP2;1 in protoplasts. We propose that Rma1H1 and Rma1 play a critical role in the downregulation of plasma membrane aquaporin levels by inhibiting aquaporin trafficking to the plasma membrane and subsequent proteasomal degradation as a response to dehydration in transgenic Arabidopsis plants.
Figures
References
-
- Alexandersson, E., Fraysse, L., Sjovall-Larsen, S., Gustavsson, S., Fellert, M., Karlsson, M., Johanson, U., and Kjellbom, P. (2005). Whole gene family expression and drought stress regulation of aquaporins. Plant Mol. Biol. 59 469–484. - PubMed
-
- Bohnert, H.J., Gong, Q., Li, P., and Ma, S. (2006). Unraveling abiotic stress tolerance mechanisms - Getting genomics going. Curr. Opin. Plant Biol. 9 180–188. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
