

The advantages of linear information processing for cerebellar computation
- PMID: 19234116
- PMCID: PMC2657437
- DOI: 10.1073/pnas.0812348106
The advantages of linear information processing for cerebellar computation
Abstract
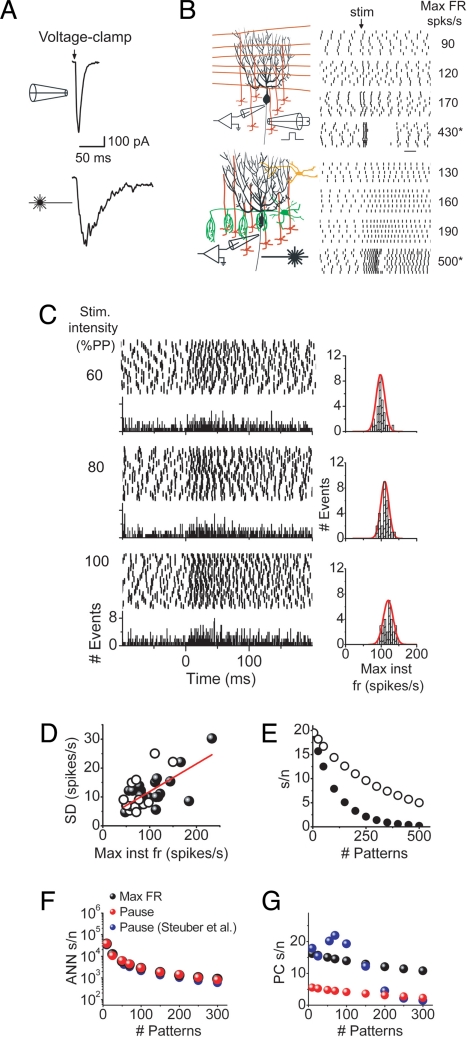
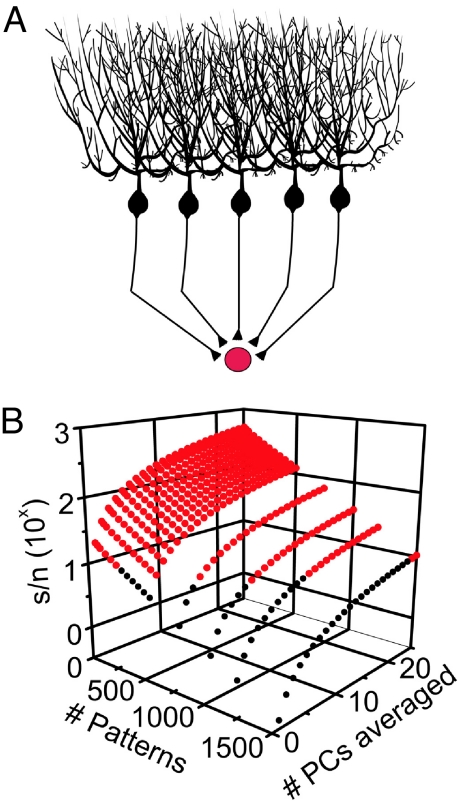
Purkinje cells can encode the strength of parallel fiber inputs in their firing by using 2 fundamentally different mechanisms, either as pauses or as linear increases in firing rate. It is not clear which of these 2 encoding mechanisms is used by the cerebellum. We used the pattern-recognition capacity of Purkinje cells based on the Marr-Albus-Ito theory of cerebellar learning to evaluate the suitability of the linear algorithm for cerebellar information processing. Here, we demonstrate the simplicity and versatility of pattern recognition in Purkinje cells linearly encoding the strength of parallel fiber inputs in their firing rate. In contrast to encoding patterns with pauses, Purkinje cells using the linear algorithm could recognize a large number of both synchronous and asynchronous input patterns in the presence or absence of inhibitory synaptic transmission. Under all conditions, the number of patterns recognized by Purkinje cells using the linear algorithm was greater than that achieved by encoding information in pauses. Linear encoding of information also allows neurons of deep cerebellar nuclei to use a simple averaging mechanism to significantly increase the computational capacity of the cerebellum. We propose that the virtues of the linear encoding mechanism make it well suited for cerebellar computation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
