

Electrochemical cues regulate assembly of the Frizzled/Dishevelled complex at the plasma membrane during planar epithelial polarization
- PMID: 19234454
- PMCID: PMC2803043
- DOI: 10.1038/ncb1836
Electrochemical cues regulate assembly of the Frizzled/Dishevelled complex at the plasma membrane during planar epithelial polarization
Erratum in
- Nat Cell Biol. 2009 Apr;11(4):508
Abstract
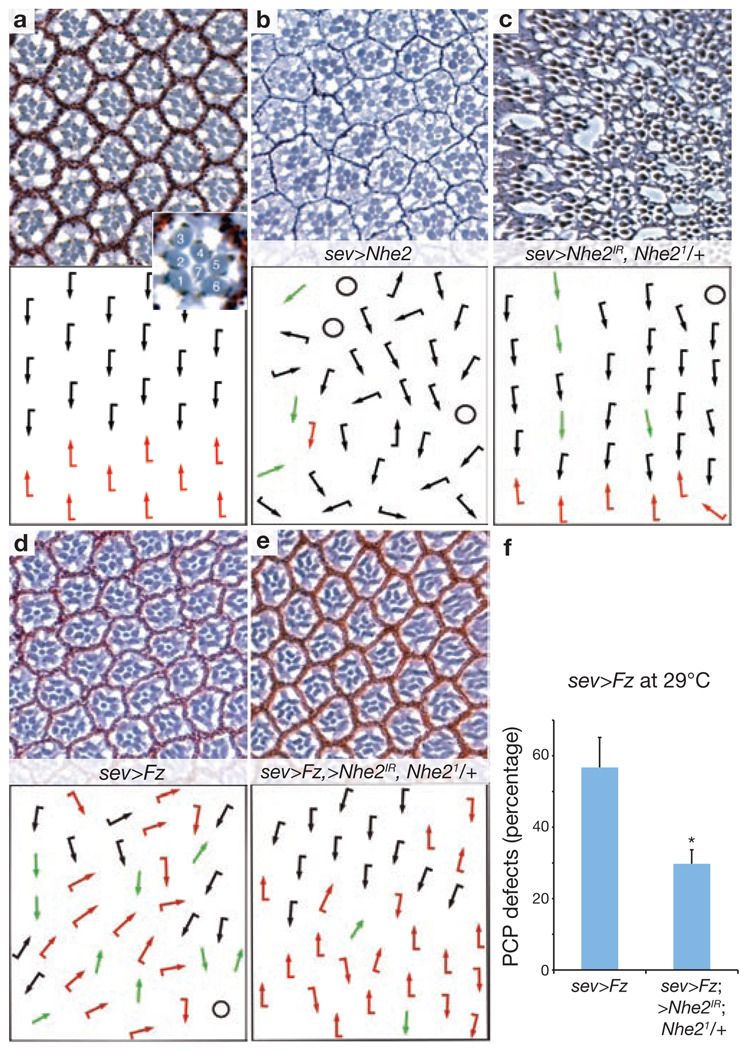
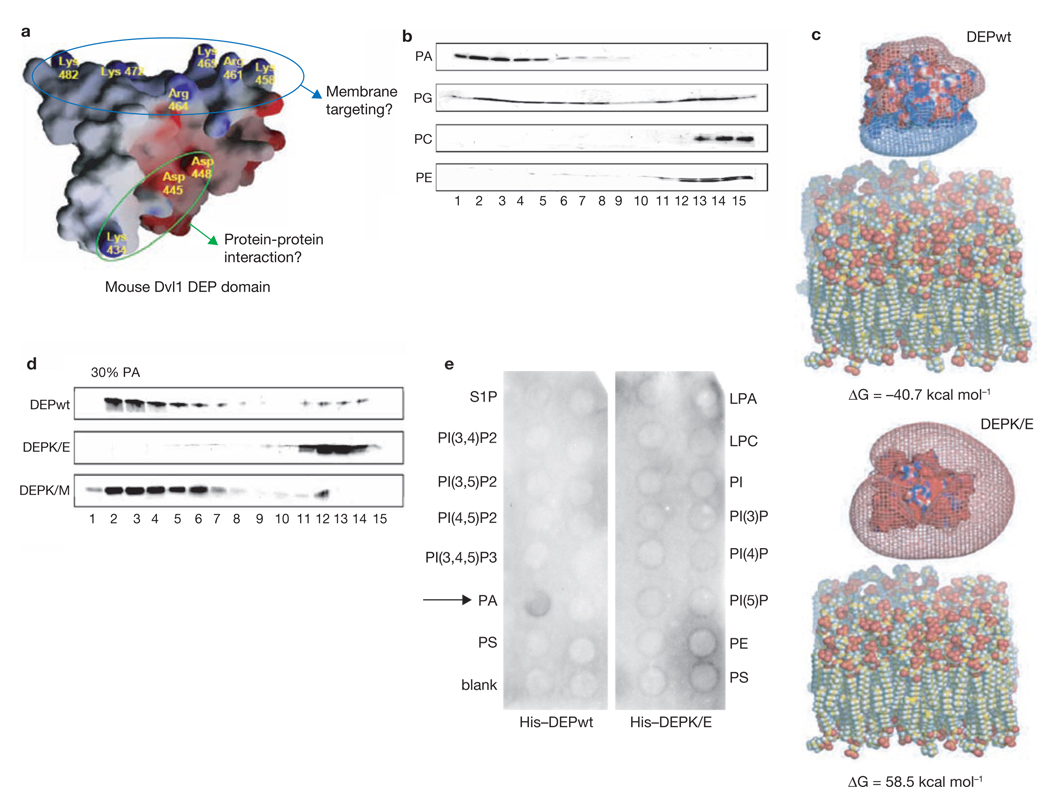
Dishevelled (Dsh) is a cytoplasmic multidomain protein that is required for all known branches of the Wnt signalling pathway. The Frizzled/planar cell polarity (Fz/PCP) signalling branch requires an asymmetric cortical localization of Dsh, but this process remains poorly understood. Using a genome-wide RNA interference (RNAi) screen in Drosophila melanogaster cells, we show that Dsh membrane localization is dependent on the Na(+)/H(+) exchange activity of the plasma membrane exchanger Nhe2. Manipulating Nhe2 expression levels in the eye causes PCP defects, and Nhe2 interacts genetically with Fz. Our data show that the binding and surface recruitment of Dsh by Fz is pH- and charge-dependent. We identify a polybasic stretch within the Dsh DEP domain that binds to negatively charged phospholipids and appears to be mechanistically important. Dsh recruitment by Fz can be abolished by converting these basic amino-acid residues into acidic ones, as in the mutant, DshKR/E. In vivo, the DshKR/E(2x) mutant with two substituted residues fails to associate with the membrane during active PCP signalling but rescues canonical Wnt signalling defects in a dsh-background. These results suggest that direct interaction between Fz and Dsh is stabilized by a pH and charge-dependent interaction of the DEP domain with phospholipids. This stabilization is particularly important for the PCP signalling branch and, thus, promotes specific pathway selection in Wnt signalling.
Figures



Comment in
-
When you are Dishevelled, fat is good and acid is bad!Nat Cell Biol. 2009 Mar;11(3):237-9. doi: 10.1038/ncb0309-237. Nat Cell Biol. 2009. PMID: 19255568
References
-
- Boutros M, Paricio N, Strutt DI, Mlodzik M. Dishevelled activates JNK and discriminates between JNK pathways in planar polarity and wingless signaling. Cell. 1998;94:109–118. - PubMed
-
- Seifert JR, Mlodzik M. Frizzled/PCP signalling: a conserved mechanism regulating cell polarity and directed motility. Nature Rev. Genet. 2007;8:126–138. - PubMed
-
- Veeman MT, Axelrod JD, Moon RT. A second canon. Functions and mechanisms of β-catenin-independent Wnt signaling. Dev. Cell. 2003;5:367–377. - PubMed
-
- Bilic J, et al. Wnt induces LRP6 signalosomes and promotes dishevelled-dependent LRP6 phosphorylation. Science. 2007;316:1619–1622. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
