

A point mutation in KINDLIN3 ablates activation of three integrin subfamilies in humans
- PMID: 19234460
- PMCID: PMC2857384
- DOI: 10.1038/nm.1917
A point mutation in KINDLIN3 ablates activation of three integrin subfamilies in humans
Abstract
Monogenic deficiency diseases provide unique opportunities to define the contributions of individual molecules to human physiology and to identify pathologies arising from their dysfunction. Here we describe a deficiency disease in two human siblings that presented with severe bleeding, frequent infections and osteopetrosis at an early age. These symptoms are consistent with but more severe than those reported for people with leukocyte adhesion deficiency III (LAD-III). Mechanistically, these symptoms arose from an inability to activate the integrins expressed on hematopoietic cells, including platelets and leukocytes. Immortalized lymphocyte cell lines isolated from the two individuals showed integrin activation defects. Several proteins previously implicated in integrin activation, including Ras-associated protein-1 (RAP1) and calcium and diacylglycerol-regulated guanine nucleotide exchange factor-1 (CALDAG-GEF1), were present and functional in these cell lines. The genetic basis for this disease was traced to a point mutation in the coding region of the KINDLIN3 (official gene symbol FERMT3) gene. When wild-type KINDLIN-3 was expressed in the immortalized lymphocytes, their integrins became responsive to activation signals. These results identify a genetic disease that severely compromises the health of the affected individuals and establish an essential role of KINDLIN-3 in integrin activation in humans. Furthermore, allogeneic bone marrow transplantation was shown to alleviate the symptoms of the disease.
Figures


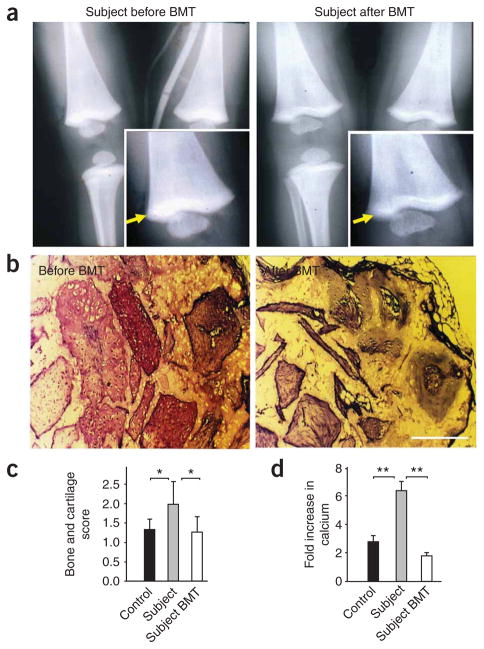
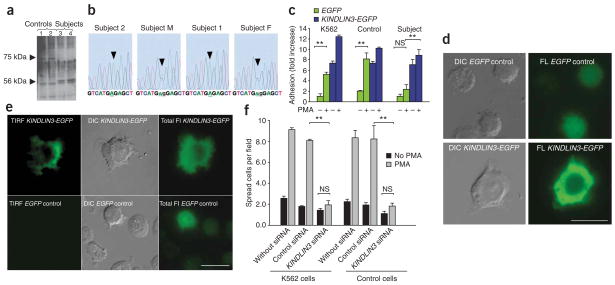
Comment in
-
When integrins fail to integrate.Nat Med. 2009 Mar;15(3):249-50. doi: 10.1038/nm0309-249. Nat Med. 2009. PMID: 19265824 Free PMC article. No abstract available.
References
-
- Hattori M, Minato N. Rap1 GTPase: functions, regulation, and malignancy. J Biochem. 2003;134:479–484. - PubMed
-
- Crittenden JR, et al. CalDAG-GEFI integrates signaling for platelet aggregation and thrombus formation. Nat Med. 2004;10:982–986. - PubMed
-
- Ussar S, Wang HV, Linder S, Fassler R, Moser M. The Kindlins: subcellular localization and expression during murine development. Exp Cell Res. 2006;312:3142–3151. - PubMed
-
- Shi X, et al. The MIG-2/integrin interaction strengthens cell-matrix adhesion and modulates cell motility. J Biol Chem. 2007;282:20455–20466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
