

Evidence that the assembly of the yeast cytochrome bc1 complex involves the formation of a large core structure in the inner mitochondrial membrane
- PMID: 19236481
- PMCID: PMC2745923
- DOI: 10.1111/j.1742-4658.2009.06916.x
Evidence that the assembly of the yeast cytochrome bc1 complex involves the formation of a large core structure in the inner mitochondrial membrane
Abstract
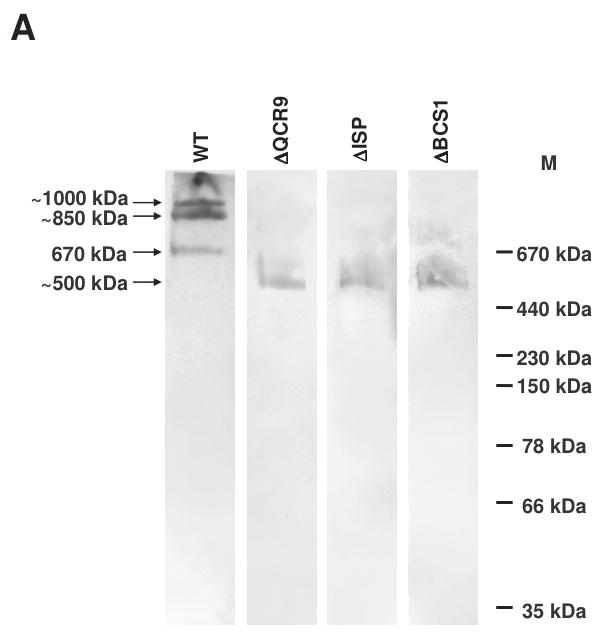
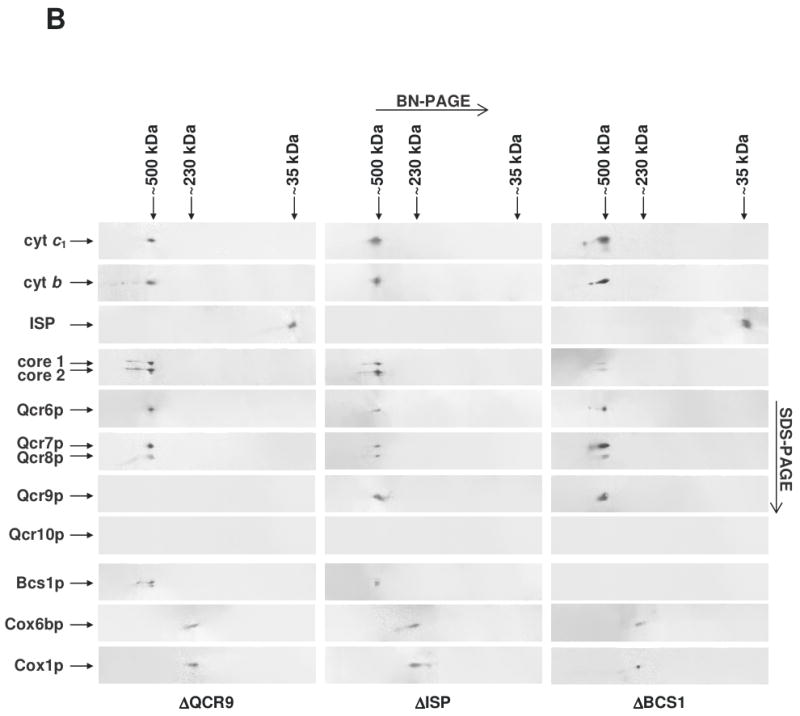
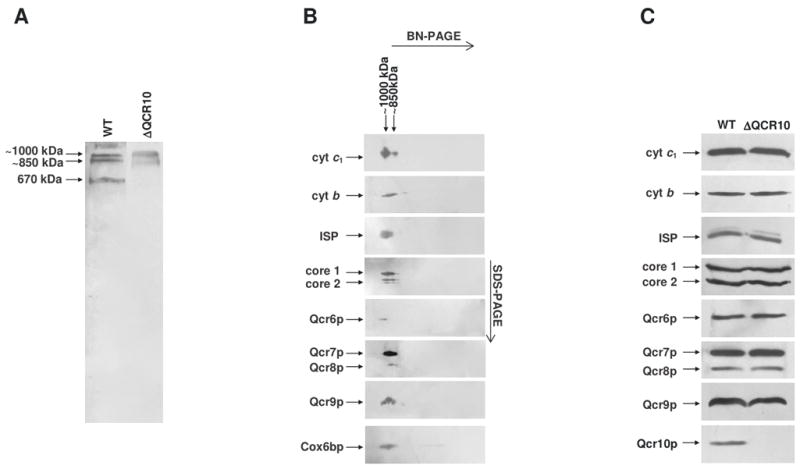
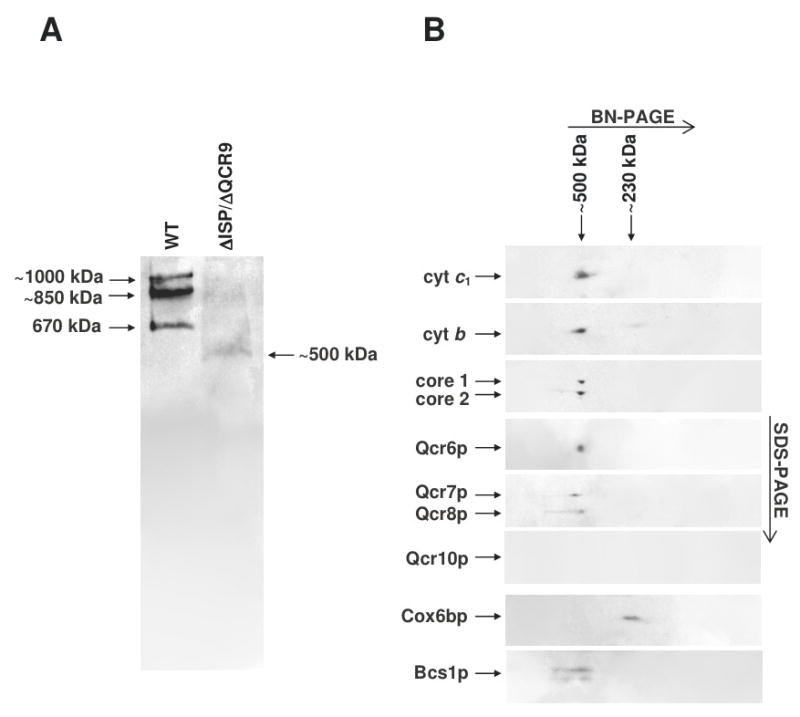
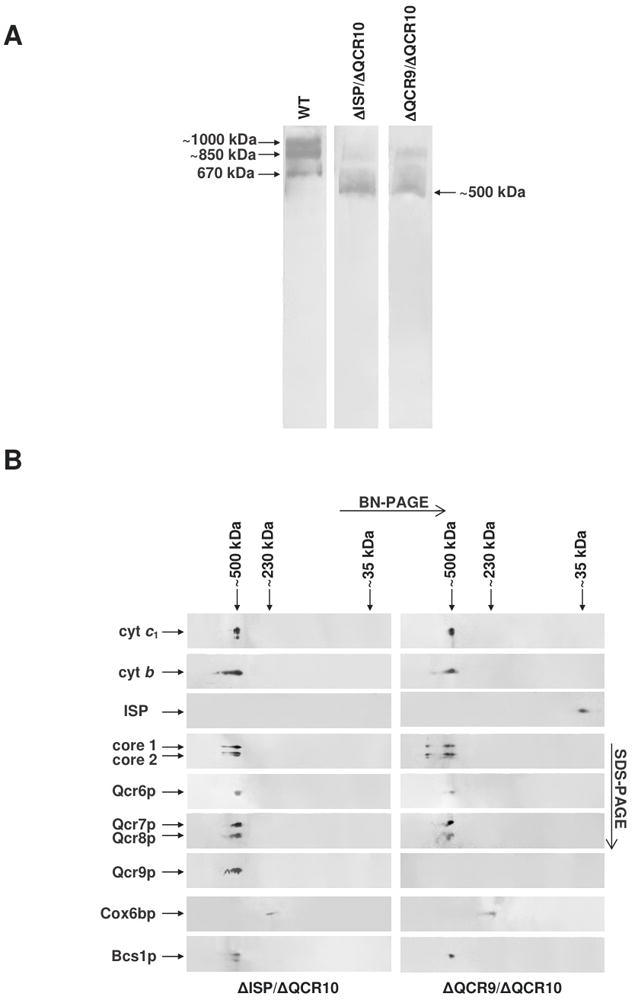
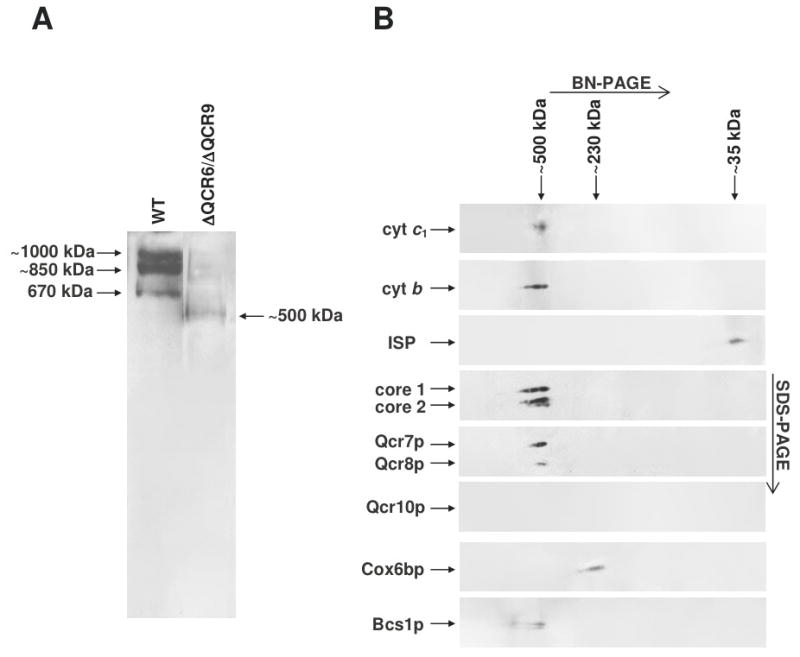
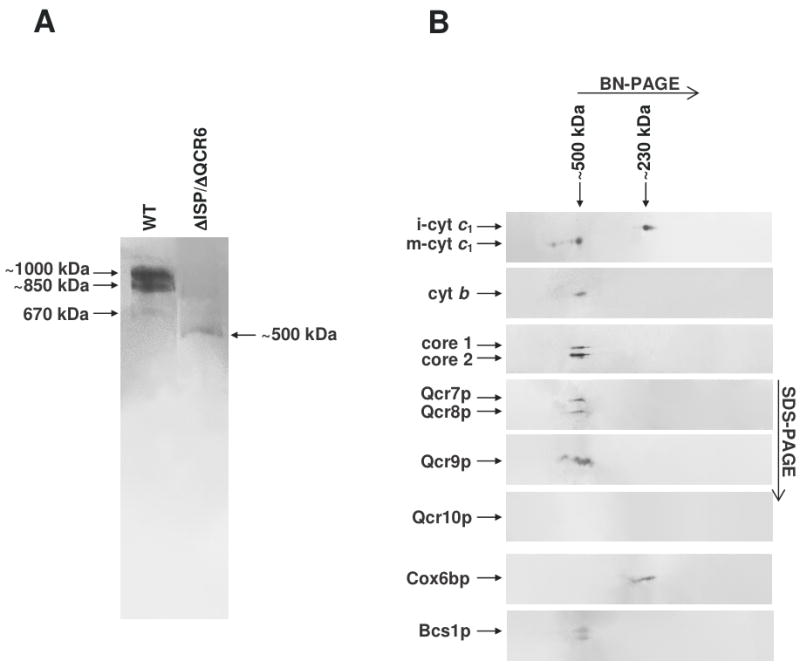
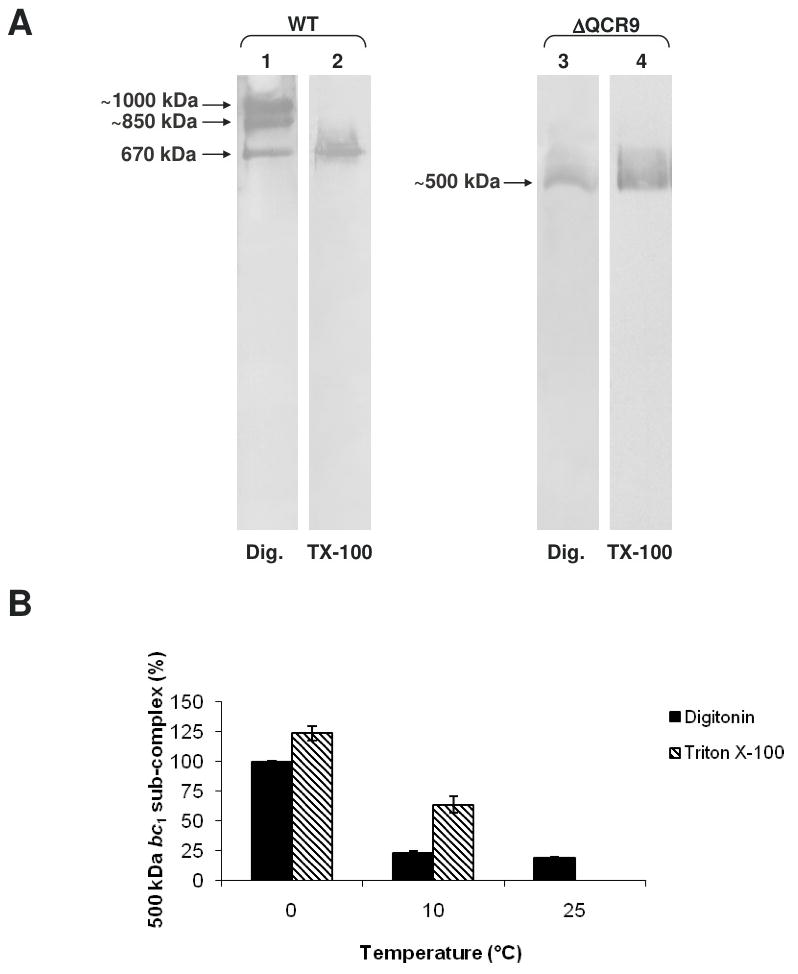
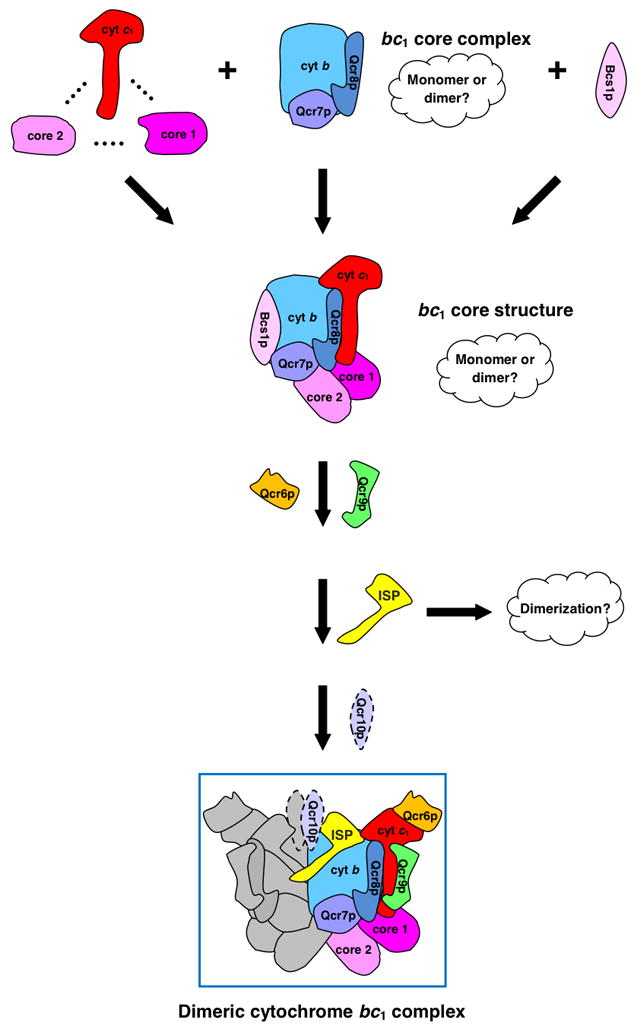
The assembly status of the cytochrome bc(1) complex has been analyzed in distinct yeast deletion strains in which genes for one or more of the bc(1) subunits were deleted. In all the yeast strains tested, a bc(1) sub-complex of approximately 500 kDa was found when the mitochondrial membranes were analyzed by blue native electrophoresis. The subsequent molecular characterization of this sub-complex, carried out in the second dimension by SDS/PAGE and immunodecoration, revealed the presence of the two catalytic subunits, cytochrome b and cytochrome c(1), associated with the noncatalytic subunits core protein 1, core protein 2, Qcr7p and Qcr8p. Together, these bc(1) subunits build up the core structure of the cytochrome bc(1) complex, which is then able to sequentially bind the remaining subunits, such as Qcr6p, Qcr9p, the Rieske iron-sulfur protein and Qcr10p. This bc(1) core structure may represent a true assembly intermediate during the maturation of the bc(1) complex; first, because of its wide distribution in distinct yeast deletion strains and, second, for its characteristics of stability, which resemble those of the intact homodimeric bc(1) complex. By contrast, the bc(1) core structure is unable to interact with the cytochrome c oxidase complex to form respiratory supercomplexes. The characterization of this novel core structure of the bc(1) complex provides a number of new elements clarifying the molecular events leading to the maturation of the yeast cytochrome bc(1) complex in the inner mitochondrial membrane.
Figures
Similar articles
-
Bcs1p can rescue a large and productive cytochrome bc(1) complex assembly intermediate in the inner membrane of yeast mitochondria.Biochim Biophys Acta. 2011 Jan;1813(1):91-101. doi: 10.1016/j.bbamcr.2010.08.004. Epub 2010 Aug 18. Biochim Biophys Acta. 2011. PMID: 20727375
-
Biogenesis of the yeast cytochrome bc1 complex.Biochim Biophys Acta. 2009 Jan;1793(1):89-96. doi: 10.1016/j.bbamcr.2008.04.011. Epub 2008 May 3. Biochim Biophys Acta. 2009. PMID: 18501197 Review.
-
Respiratory complex III dysfunction in humans and the use of yeast as a model organism to study mitochondrial myopathy and associated diseases.Biochim Biophys Acta. 2013 Nov-Dec;1827(11-12):1346-61. doi: 10.1016/j.bbabio.2012.11.015. Epub 2012 Dec 5. Biochim Biophys Acta. 2013. PMID: 23220121 Review.
-
Further insights into the assembly of the yeast cytochrome bc1 complex based on analysis of single and double deletion mutants lacking supernumerary subunits and cytochrome b.Eur J Biochem. 2004 Mar;271(6):1209-18. doi: 10.1111/j.1432-1033.2004.04024.x. Eur J Biochem. 2004. PMID: 15009199
-
The N-terminus of the Qcr7 protein of the cytochrome bc(1) complex in S. cerevisiae may be involved in facilitating stability of the subcomplex with the Qcr8 protein and cytochrome b.Arch Biochem Biophys. 2001 Sep 15;393(2):215-21. doi: 10.1006/abbi.2001.2498. Arch Biochem Biophys. 2001. PMID: 11556808
Cited by
-
Late-stage maturation of the Rieske Fe/S protein: Mzm1 stabilizes Rip1 but does not facilitate its translocation by the AAA ATPase Bcs1.Mol Cell Biol. 2012 Nov;32(21):4400-9. doi: 10.1128/MCB.00441-12. Epub 2012 Aug 27. Mol Cell Biol. 2012. PMID: 22927643 Free PMC article.
-
The LYR protein Mzm1 functions in the insertion of the Rieske Fe/S protein in yeast mitochondria.Mol Cell Biol. 2011 Oct;31(19):3988-96. doi: 10.1128/MCB.05673-11. Epub 2011 Aug 1. Mol Cell Biol. 2011. PMID: 21807901 Free PMC article.
-
Coordination of cytochrome bc1 complex assembly at MICOS.EMBO Rep. 2025 Jan;26(2):353-384. doi: 10.1038/s44319-024-00336-x. Epub 2024 Dec 2. EMBO Rep. 2025. PMID: 39623166 Free PMC article.
-
Structure, function, and assembly of heme centers in mitochondrial respiratory complexes.Biochim Biophys Acta. 2012 Sep;1823(9):1604-16. doi: 10.1016/j.bbamcr.2012.04.008. Epub 2012 Apr 24. Biochim Biophys Acta. 2012. PMID: 22554985 Free PMC article. Review.
-
Assembly of the Multi-Subunit Cytochrome bc1 Complex in the Yeast Saccharomyces cerevisiae.Int J Mol Sci. 2022 Sep 11;23(18):10537. doi: 10.3390/ijms231810537. Int J Mol Sci. 2022. PMID: 36142449 Free PMC article. Review.
References
-
- Trumpower BL. The cytochrome bc1 Complex. In: Lennarz V, Lane MD, editors. Encyclopedia of Biological Chemistry. Elsevier Inc; Amsterdam: 2004. pp. 528–534.
-
- Smith JL, Zhang H, Yan J, Kurisuand G, Cramer WA. Cytochrome bc complexes: a common core of structure and function surrounded by diversity in the outlying provinces. Curr Op Struct Biol. 2004;14:432–439. - PubMed
-
- Zara V, Conte L, Trumpower BL. Biogenesis of the yeast cytochrome bc1 complex. Biochim Biophys Acta. 2009;1793:89–96. - PubMed
-
- Hunte C, Solmaz S, Palsdottir H, Wenz T. A structural perspective on mechanism and function of the cytochrome bc1 complex. Results Prob Cell Differ. 2008;45:253–278. - PubMed
-
- Xia D, Yu C-A, Kim H, Xia J-Z, Kachurin AM, Zhang L, Yu L, Deinsenhofer J. Crystal structure of the cytochrome bc1 complex of bovine heart mitochondria. Science. 1997;277:60–66. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
