

Cadmium-induced testicular injury
- PMID: 19236889
- PMCID: PMC2804910
- DOI: 10.1016/j.taap.2009.01.028
Cadmium-induced testicular injury
Abstract
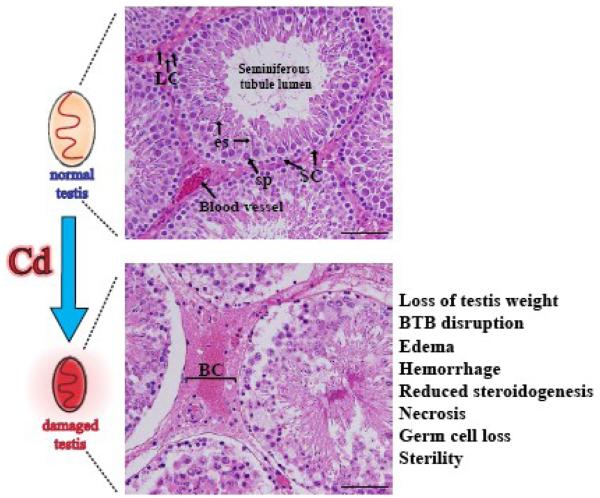
Cadmium (Cd) is an environmental toxicant and an endocrine disruptor in humans and rodents. Several organs (e.g., kidney, liver) are affected by Cd and recent studies have illustrated that the testis is exceedingly sensitive to Cd toxicity. More important, Cd and other toxicants, such as heavy metals (e.g., lead, mercury) and estrogenic-based compounds (e.g., bisphenols) may account for the recent declining fertility in men among developed countries by reducing sperm count and testis function. In this review, we critically discuss recent data in the field that have demonstrated the Cd-induced toxicity to the testis is probably the result of interactions of a complex network of causes. This is likely to involve the disruption of the blood-testis barrier (BTB) via specific signal transduction pathways and signaling molecules, such as p38 mitogen-activated protein kinase (MAPK). We also summarize current studies on factors that confer and/or regulate the testis sensitivity to Cd, such as Cd transporters and metallothioneins, the impact of Cd on the testis as an endocrine disruptor and oxidative stress inducer, and how it may disrupt the Zn(2+) and/or Ca(2+) mediated cellular events. While much work is needed before a unified mechanistic pathway of Cd-induced testicular toxicity emerges, recent studies have helped to identify some of the likely mechanisms and/or events that take place during Cd-induced testis injury. Furthermore, some of the recent studies have shed lights on potential therapeutic or preventive approaches that can be developed in future studies by blocking or minimizing the destructive effects of Cd to testicular function in men.
Figures



References
-
- Acharya UR, Mishra M, Patro J, Panda MK. Effect of vitamins C and E on spermatogenesis in mice exposed to cadmium. Reprod. Toxicol. 2008;25:84–88. - PubMed
-
- Amara S, Abdelmelek H, Garrel C, Guiraud P, Douki T, Ravanat JL, Favier A, Sakly M, Ben Rhouma K. Preventive effect of zinc against cadmium-induced oxidative stress in the rat testis. J. Reprod. Dev. 2008;54:129–134. - PubMed
-
- ATSDR . Agency for Toxic Substances and Disease Registry. U.S. Department of Health and Human Services; Atlanta, GA: 2007. The 2007 CERCLA priority list of hazardous substances.
-
- ATSDR . Agency for Toxic Substances and Disease Registry. U.S. Department of Health and Human Services; Atlanta, GA: 2008. Cadmium toxicity - Case Studies in Environmental Medicine.
-
- Beardsley A, Robertson DM, O'Donnell L. A complex containing a6b1-integrin and phosphorylated focal adhesion kinase between Sertoli cells and elongated spermatids during spermatid release from the seminiferous epithelium. J. Endocrinol. 2006;190:759–770. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
