

Peroxisome proliferator-activated receptor gamma activation restores islet function in diabetic mice through reduction of endoplasmic reticulum stress and maintenance of euchromatin structure
- PMID: 19237535
- PMCID: PMC2663298
- DOI: 10.1128/MCB.01179-08
Peroxisome proliferator-activated receptor gamma activation restores islet function in diabetic mice through reduction of endoplasmic reticulum stress and maintenance of euchromatin structure
Abstract
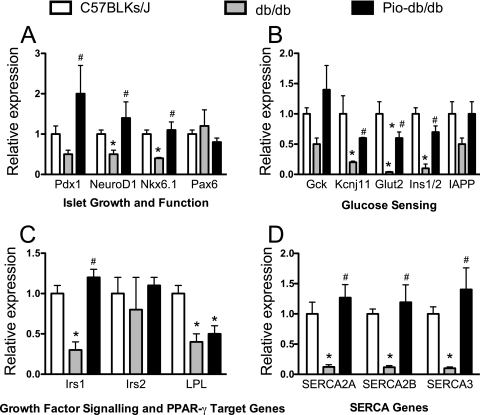
The nuclear receptor peroxisome proliferator-activated receptor gamma (PPAR-gamma) is an important target in diabetes therapy, but its direct role, if any, in the restoration of islet function has remained controversial. To identify potential molecular mechanisms of PPAR-gamma in the islet, we treated diabetic or glucose-intolerant mice with the PPAR-gamma agonist pioglitazone or with a control. Treated mice exhibited significantly improved glycemic control, corresponding to increased serum insulin and enhanced glucose-stimulated insulin release and Ca(2+) responses from isolated islets in vitro. This improved islet function was at least partially attributed to significant upregulation of the islet genes Irs1, SERCA, Ins1/2, and Glut2 in treated animals. The restoration of the Ins1/2 and Glut2 genes corresponded to a two- to threefold increase in the euchromatin marker histone H3 dimethyl-Lys4 at their respective promoters and was coincident with increased nuclear occupancy of the islet methyltransferase Set7/9. Analysis of diabetic islets in vitro suggested that these effects resulting from the presence of the PPAR-gamma agonist may be secondary to improvements in endoplasmic reticulum stress. Consistent with this possibility, incubation of thapsigargin-treated INS-1 beta cells with the PPAR-gamma agonist resulted in the reduction of endoplasmic reticulum stress and restoration of Pdx1 protein levels and Set7/9 nuclear occupancy. We conclude that PPAR-gamma agonists exert a direct effect in diabetic islets to reduce endoplasmic reticulum stress and enhance Pdx1 levels, leading to favorable alterations of the islet gene chromatin architecture.
Figures














References
-
- Amrani, A., S. Durant, M. Throsby, J. Coulaud, M. Dardenne, and F. Homo-Delarche. 1998. Glucose homeostasis in the nonobese diabetic mouse at the prediabetic stage. Endocrinology 1391115-1124. - PubMed
-
- Brunham, L. R., J. K. Kruit, T. D. Pape, J. M. Timmins, A. Q. Reuwer, Z. Vasanji, B. J. Marsh, B. Rodrigues, J. D. Johnson, J. S. Parks, C. B. Verchere, and M. R. Hayden. 2007. Beta-cell ABCA1 influences insulin secretion, glucose homeostasis and response to thiazolidinedione treatment. Nat. Med. 13340-347. - PubMed
-
- Campbell, I. W., and S. Mariz. 2007. Beta-cell preservation with thiazolidinediones. Diabetes Res. Clin. Pract. 76163-176. - PubMed
-
- Chakrabarti, S. K., J. Francis, S. M. Ziesmann, J. C. Garmey, and R. G. Mirmira. 2003. Covalent histone modifications underlie the developmental regulation of insulin gene transcription in pancreatic beta cells. J. Biol. Chem. 27823617-23623. - PubMed
-
- Chakrabarti, S. K., J. C. James, and R. G. Mirmira. 2002. Quantitative assessment of gene targeting in vitro and in vivo by the pancreatic transcription factor, Pdx1. Importance of chromatin structure in directing promoter binding. J. Biol. Chem. 27713286-13293. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous