

Reactive oxygen species facilitate adipocyte differentiation by accelerating mitotic clonal expansion
- PMID: 19237544
- PMCID: PMC2667747
- DOI: 10.1074/jbc.M808742200
Reactive oxygen species facilitate adipocyte differentiation by accelerating mitotic clonal expansion
Abstract
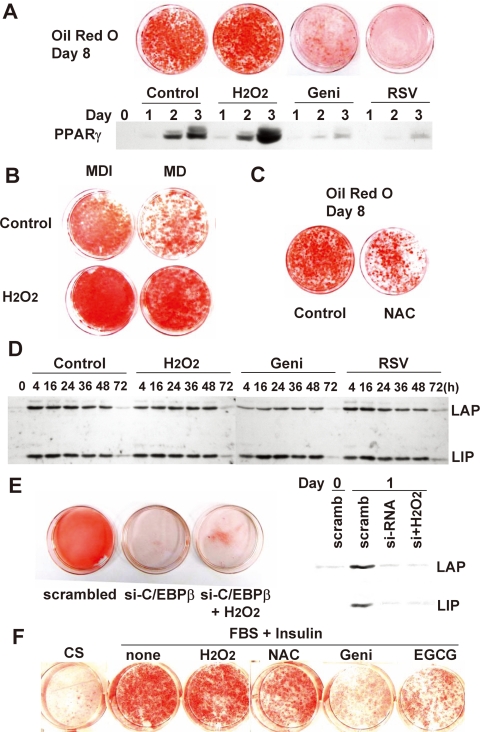
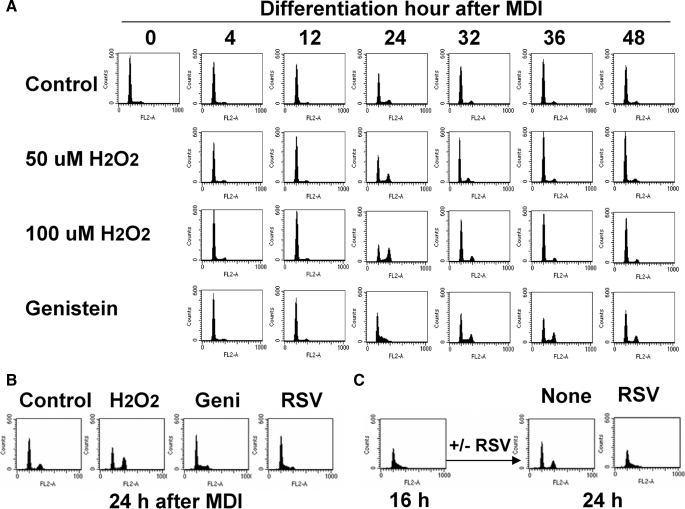
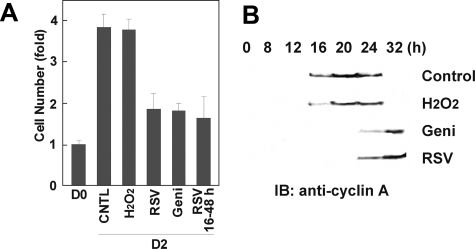
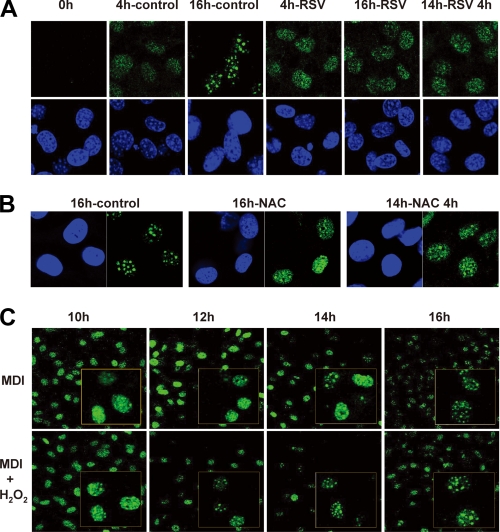
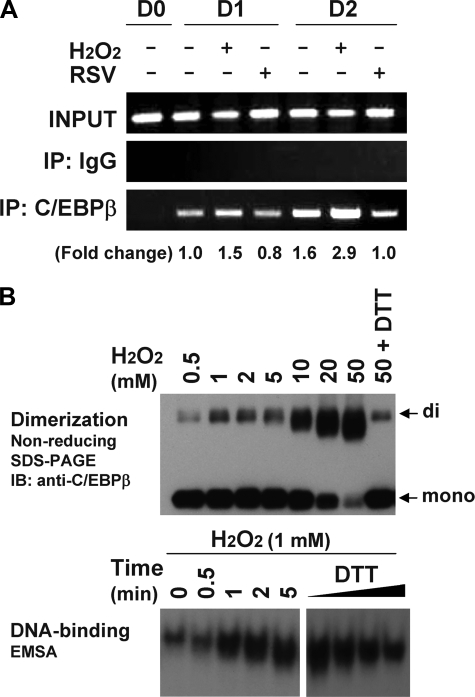
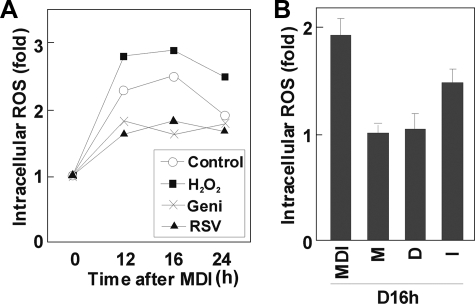
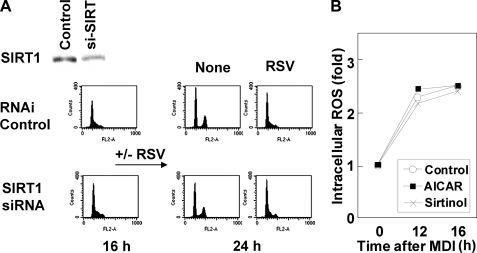
Growth-arrested 3T3-L1 preadipocytes rapidly express CCAAT/enhancer-binding protein-beta (C/EBPbeta) upon hormonal induction of differentiation. However, the DNA binding activity of C/EBPbeta is not activated until the cells synchronously reenter S phase during the mitotic clonal expansion (MCE) phase of differentiation. In this period, C/EBPbeta is sequentially phosphorylated by MAPK and glycogen synthase kinase-3beta, inducing C/EBPbeta DNA binding activity and transcription of its target genes. Because the DNA binding activity of C/EBPbeta is further enhanced by oxidation in vitro, we investigated how redox state affects C/EBPbeta DNA binding and MCE during adipogenesis. When 3T3-L1 cells were treated with H(2)O(2) and hormonal stimuli, differentiation was accelerated with increased expression of peroxisome proliferator-activated receptor gamma. Interestingly, cell cycle progression (S to G(2)/M phase) was markedly enhanced by H(2)O(2), whereas antioxidants caused an S phase arrest during the MCE. H(2)O(2) treatment resulted in the early appearance of a punctate pattern observed by immunofluorescent staining of C/EBPbeta, which is a hallmark for C/EBPbeta binding to regulatory elements, whereas a short antioxidant treatment rapidly dispersed the centromeric localization of C/EBPbeta. Consistently, reactive oxygen species production was increased during 3T3-L1 differentiation. Our results indicate that redox-induced C/EBPbeta DNA binding activity, along with the dual phosphorylation of C/EBPbeta, is required for the MCE and terminal differentiation of adipocytes.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
