

Glycogen synthase kinase 3beta missplicing contributes to leukemia stem cell generation
- PMID: 19237556
- PMCID: PMC2646624
- DOI: 10.1073/pnas.0900189106
Glycogen synthase kinase 3beta missplicing contributes to leukemia stem cell generation
Abstract
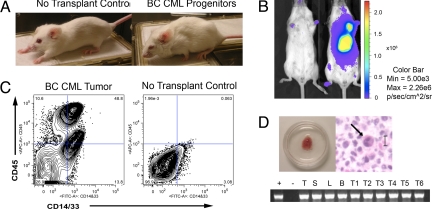
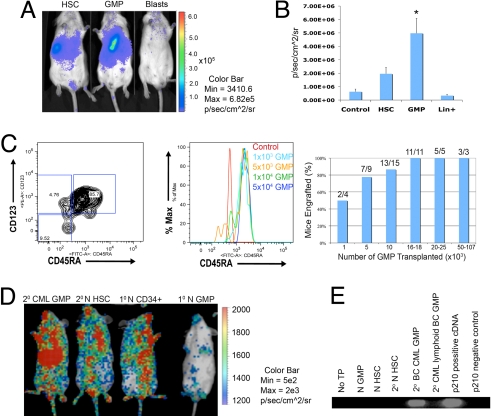
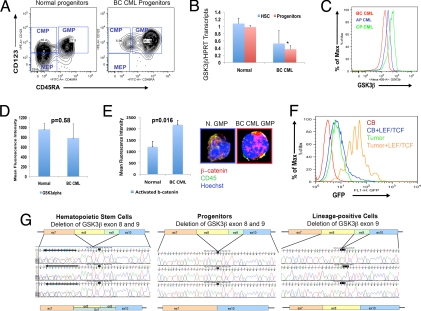
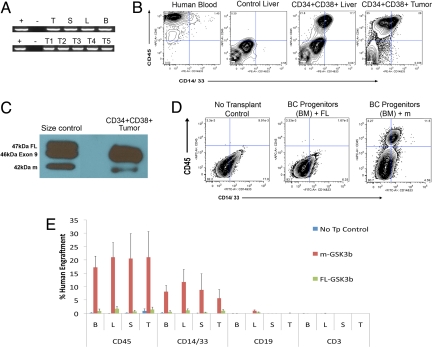
Recent evidence suggests that a rare population of self-renewing cancer stem cells (CSC) is responsible for cancer progression and therapeutic resistance. Chronic myeloid leukemia (CML) represents an important paradigm for understanding the genetic and epigenetic events involved in CSC production. CML progresses from a chronic phase (CP) in hematopoietic stem cells (HSC) that harbor the BCR-ABL translocation, to blast crisis (BC), characterized by aberrant activation of beta-catenin within granulocyte-macrophage progenitors (GMP). A major barrier to predicting and inhibiting blast crisis transformation has been the identification of mechanisms driving beta-catenin activation. Here we show that BC CML myeloid progenitors, in particular GMP, serially transplant leukemia in immunocompromised mice and thus are enriched for leukemia stem cells (LSC). Notably, cDNA sequencing of Wnt/beta-catenin pathway regulatory genes, including adenomatous polyposis coli, GSK3beta, axin 1, beta-catenin, lymphoid enhancer factor-1, cyclin D1, and c-myc, revealed a novel in-frame splice deletion of the GSK3beta kinase domain in the GMP of BC samples that was not detectable by sequencing in blasts or normal progenitors. Moreover, BC CML progenitors with misspliced GSK3beta have enhanced beta-catenin expression as well as serial engraftment potential while reintroduction of full-length GSK3beta reduces both in vitro replating and leukemic engraftment. We propose that CP CML is initiated by BCR-ABL expression in an HSC clone but that progression to BC may include missplicing of GSK3beta in GMP LSC, enabling unphosphorylated beta-catenin to participate in LSC self-renewal. Missplicing of GSK3beta represents a unique mechanism for the emergence of BC CML LSC and might provide a novel diagnostic and therapeutic target.
Conflict of interest statement
Conflict of interest statement: I.L.W. has equity ownership in Cellerant, Inc. and Stemcells, Inc. C.H.M.J consults for Wintherix Inc.
Figures
References
-
- Jamieson CH, et al. Granulocyte-macrophage progenitors as candidate leukemic stem cells in blast-crisis CML. N Engl J Med. 2004;351:657–667. - PubMed
-
- Daley GQ, Van Etten RA, Baltimore D. Induction of chronic myelogenous leukemia in mice by the P210bcr/abl gene of the Philadelphia chromosome. Science. 1990;247:824–830. - PubMed
-
- Druker BJ, et al. Five-year follow-up of patients receiving imatinib for chronic myeloid leukemia. N Engl J Med. 2006;355:2408–2417. - PubMed
-
- Fialkow PJ, Jacobson RJ, Papayannopoulou T. Chronic myelocytic leukemia: Clonal origin in a stem cell common to the granulocyte, erythrocyte, platelet, and monocyte/macrophage. Am J Med. 1977;63:125–130. - PubMed
-
- Hungerford DA. The Philadelphia Chromosome and Some Others. Ann Intern Med. 1964;61:789–793. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
