

Interaction between the N- and C-terminal domains modulates the stability and lipid binding of apolipoprotein A-I
- PMID: 19239199
- PMCID: PMC2936823
- DOI: 10.1021/bi802317v
Interaction between the N- and C-terminal domains modulates the stability and lipid binding of apolipoprotein A-I
Abstract
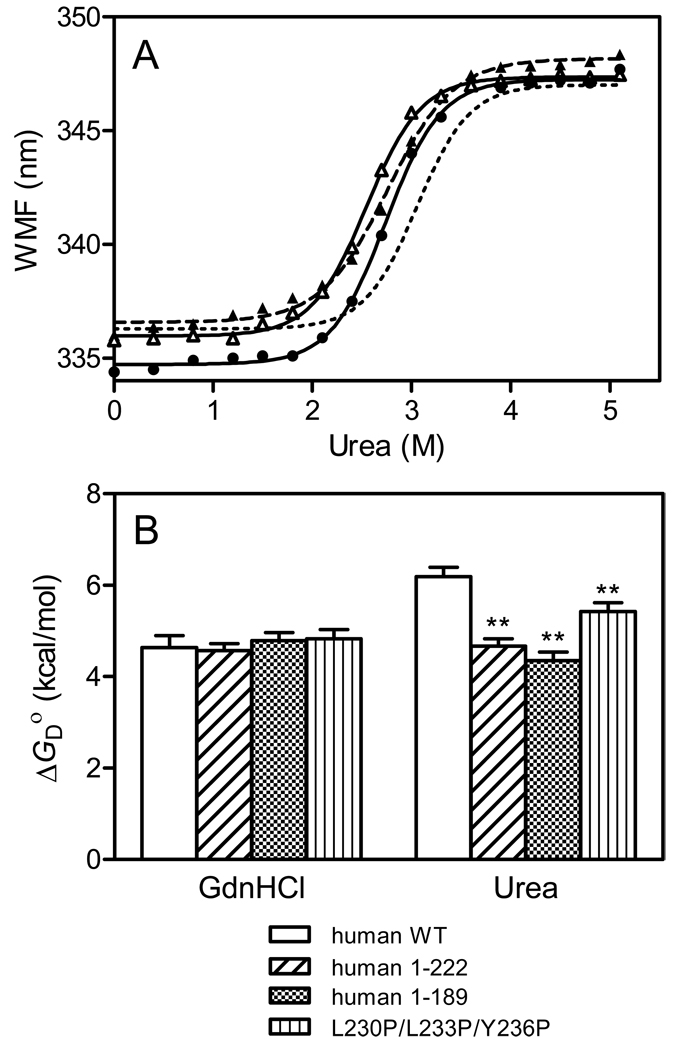
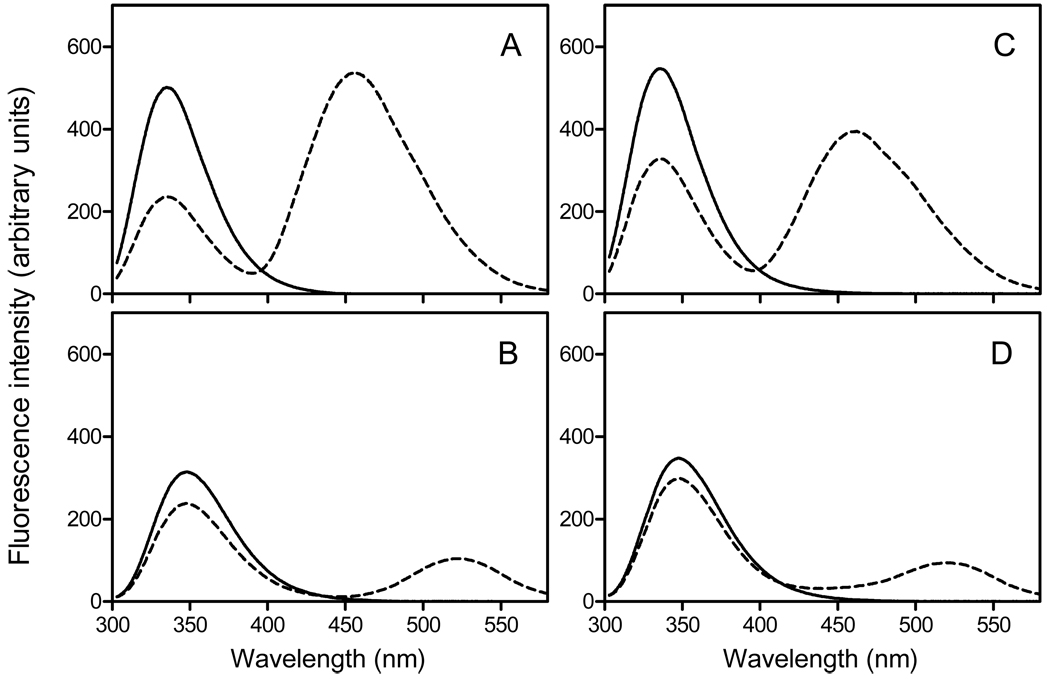
The tertiary structures of human and mouse apolipoprotein A-I (apoA-I) are comprised of an N-terminal helix bundle and a separate C-terminal domain. To define the possible intramolecular interaction between the N- and the C-terminal domains, we examined the effects on protein stability and lipid-binding properties of exchanging either the C-terminal domain or helix between human and mouse apoA-I. Chemical denaturation experiments demonstrated that replacement of the C-terminal domain or helical segment in human apoA-I with the mouse counterparts largely destabilizes the N-terminal helix bundle. Removal of the C-terminal domain or alpha-helix in human apoA-I had a similar effect on the destabilization of the helix bundle against urea denaturation, indicating that the C-terminal helical segment mainly contributes to stabilizing the N-terminal helix bundle structure in the apoA-I molecule. Consistent with this, KI quenching experiments indicated that removal or replacement of the C-terminal domain or helix in human apoA-I causes Trp residues in the N-terminal domain to become exposed to solvent. Measurements of the heats of binding to egg phosphatidylcholine (PC) vesicles and the kinetics of solubilization of dimyristoyl PC vesicles demonstrated that the destabilized human N-terminal helix bundle can strongly interact with lipids without the hydrophobic C-terminal helix. In addition, site-specific labeling of the N- and C-terminal helices by acrylodan to probe the conformational stability and the spatial proximity of the two domains indicated that the C-terminal helix is located near the N-terminal helix bundle, leading to a relatively less solvent-exposed, more organized conformation of the C-terminal domain. Taken together, these results suggest that interaction between the N- and C-terminal tertiary structure domains in apoA-I modulates the stability and lipid-binding properties of the N-terminal helix bundle.
Figures



References
-
- Curtiss LK, Valenta DT, Hime NJ, Rye KA. What is so special about apolipoprotein AI in reverse cholesterol transport? Arterioscler Thromb Vasc Biol. 2006;26:12–19. - PubMed
-
- Tall AR, Yvan-Charvet L, Terasaka N, Pagler T, Wang N. HDL, ABC transporters, and cholesterol efflux: implications for the treatment of atherosclerosis. Cell Metab. 2008;7:365–375. - PubMed
-
- Krimbou L, Marcil M, Genest J. New insights into the biogenesis of human high-density lipoproteins. Curr Opin Lipidol. 2006;17:258–267. - PubMed
-
- Oram JF, Vaughan AM. ATP-Binding cassette cholesterol transporters and cardiovascular disease. Circ Res. 2006;99:1031–1043. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
