

Dissection of kinesin's processivity
- PMID: 19242550
- PMCID: PMC2644789
- DOI: 10.1371/journal.pone.0004612
Dissection of kinesin's processivity
Abstract
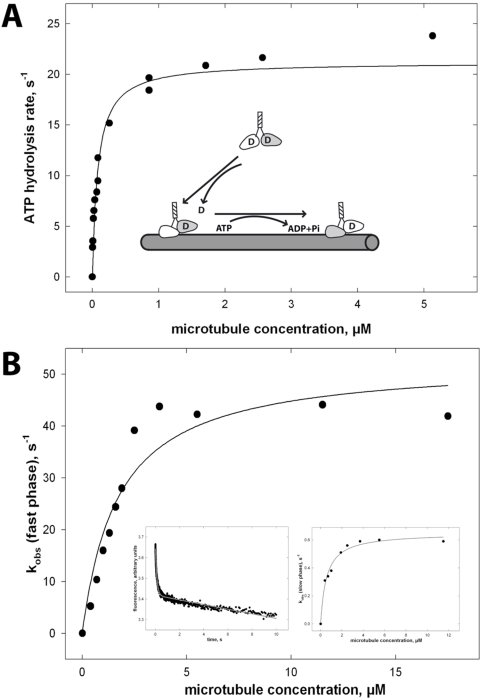
The protein family of kinesins contains processive motor proteins that move stepwise along microtubules. This mechanism requires the precise coupling of the catalytic steps in the two heads, and their precise mechanical coordination. Here we show that these functionalities can be uncoupled in chimera of processive and non-processive kinesins. A chimera with the motor domain of Kinesin-1 and the dimerization domain of a non-processive Kinesin-3 motor behaves qualitatively as conventional kinesin and moves processively in TIRF and bead motility assays, suggesting that spatial proximity of two Kinein-1 motor domains is sufficient for processive behavior. In the reverse chimera, the non-processive motor domains are unable to step along microtubules, despite the presence of the Kinesin-1 neck coiled coil. Still, ATP-binding to one head of these chimera induces ADP-release from the partner head, a characteristic feature of alternating site catalysis. These results show that processive movement of kinesin dimers requires elements in the motor head that respond to ADP-release and induce stepping, in addition to a proper spacing of the motor heads via the neck coiled coil.
Conflict of interest statement
Figures





Similar articles
-
Role of the kinesin neck region in processive microtubule-based motility.J Cell Biol. 1998 Mar 23;140(6):1407-16. doi: 10.1083/jcb.140.6.1407. J Cell Biol. 1998. PMID: 9508773 Free PMC article.
-
Highly processive microtubule-stimulated ATP hydrolysis by dimeric kinesin head domains.Nature. 1995 Oct 5;377(6548):448-50. doi: 10.1038/377448a0. Nature. 1995. PMID: 7566125
-
Loading direction regulates the affinity of ADP for kinesin.Nat Struct Biol. 2003 Apr;10(4):308-11. doi: 10.1038/nsb911. Nat Struct Biol. 2003. PMID: 12640444
-
Kinesin, 30 years later: Recent insights from structural studies.Protein Sci. 2015 Jul;24(7):1047-56. doi: 10.1002/pro.2697. Epub 2015 Jun 11. Protein Sci. 2015. PMID: 25975756 Free PMC article. Review.
-
Kinesin: a molecular motor with a spring in its step.Proc Biol Sci. 2002 Nov 22;269(1507):2363-71. doi: 10.1098/rspb.2002.2117. Proc Biol Sci. 2002. PMID: 12495505 Free PMC article. Review.
Cited by
-
These motors were made for walking.Protein Sci. 2020 Aug;29(8):1707-1723. doi: 10.1002/pro.3895. Epub 2020 Jun 26. Protein Sci. 2020. PMID: 32472639 Free PMC article. Review.
-
Drosophila PAT1 is required for Kinesin-1 to transport cargo and to maximize its motility.Development. 2010 Aug;137(16):2763-72. doi: 10.1242/dev.048108. Epub 2010 Jul 14. Development. 2010. PMID: 20630947 Free PMC article.
-
FRET measurements of kinesin neck orientation reveal a structural basis for processivity and asymmetry.Proc Natl Acad Sci U S A. 2010 Mar 23;107(12):5453-8. doi: 10.1073/pnas.0914924107. Epub 2010 Mar 8. Proc Natl Acad Sci U S A. 2010. PMID: 20212149 Free PMC article.
-
Covalent immobilization of microtubules on glass surfaces for molecular motor force measurements and other single-molecule assays.Methods Mol Biol. 2014;1136:137-69. doi: 10.1007/978-1-4939-0329-0_9. Methods Mol Biol. 2014. PMID: 24633798 Free PMC article.
-
Detailed tuning of structure and intramolecular communication are dispensable for processive motion of myosin VI.Biophys J. 2011 Jan 19;100(2):430-9. doi: 10.1016/j.bpj.2010.11.045. Biophys J. 2011. PMID: 21244839 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
