

Review
doi: 10.1021/cr800544v.
Protein quality control as a strategy for cellular regulation: lessons from ubiquitin-mediated regulation of the sterol pathway
Affiliations
- PMID: 19243134
- PMCID: PMC8530524
- DOI: 10.1021/cr800544v
Item in Clipboard
Review
Protein quality control as a strategy for cellular regulation: lessons from ubiquitin-mediated regulation of the sterol pathway
Chem Rev.
2009 Apr.
No abstract available
Figures
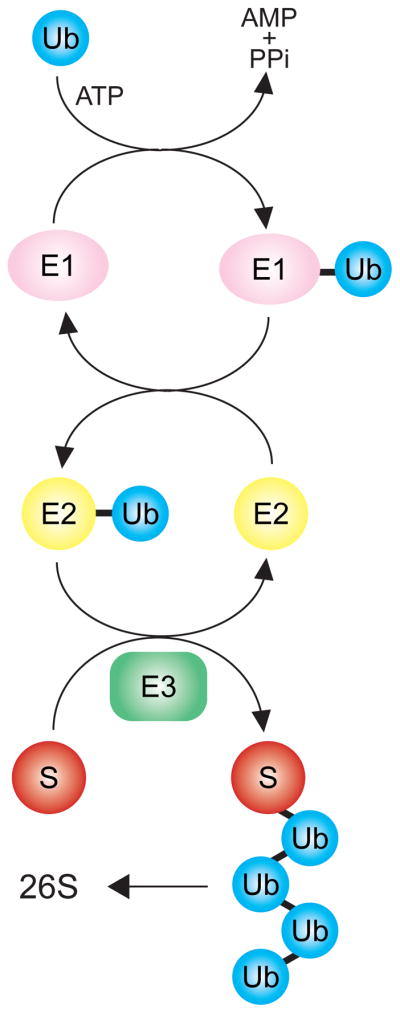
The covalent attachment of the small (7.6 kD) protein ubiquitin (Ub) to a target substrate (S) proceeds by a cascade of enzymes. The ubiquitin activating enzyme (UBE), or E1, adds Ub to itself in a thioester linkage on the E1. Ubiquitin conjugating enzymes (UBCs), E2s, are charged by transfer of ubiquitin from the E1 to a thioester linkage on the E2. Finally, a ubiquitin ligase, or E3, brokers the transfer of ubiquitin to the target substrate S, producing a ubiquitinated protein. This process is continued by continued addition of the next ubiquitin to the previously added one, producing a polyubiquitin chain. The polyubiquitin chain is recognized with high specificity by the 26S proteasome, causing degradation of the S target. The E3 is thus a critical determininat of specificity in this modification scheme. Typically, there is one E1, tens of E2s (10 in yeast, ~ 50 in mammals) and many E3s.
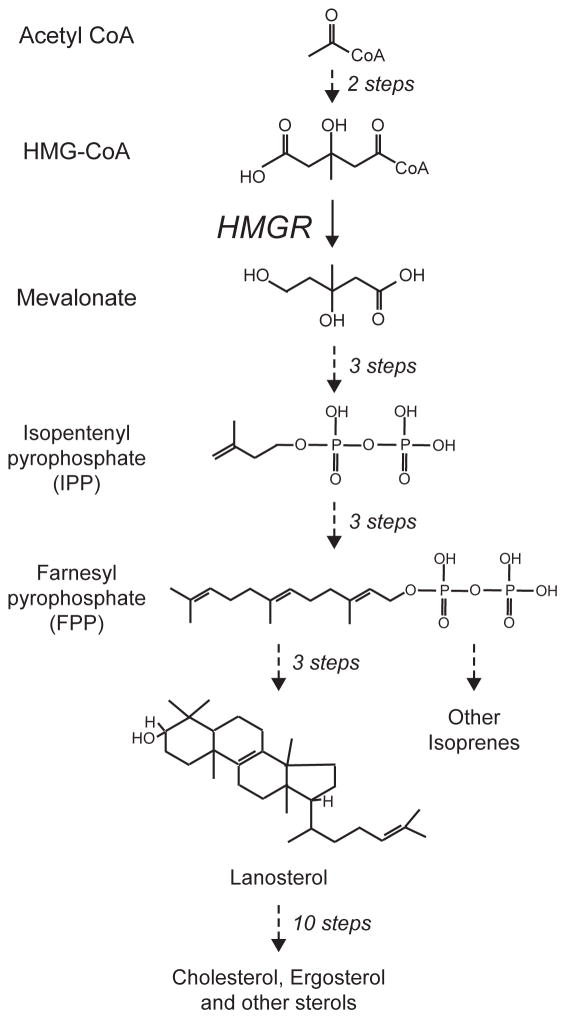
The synthesis of sterols and other isoprenes occurs by the sequential construction of the 5 carbon pyrophosphorylated isoprenes IPP and its isomer DMAPP (not shown), followed by iterated condensation to produce 10, 15 and higher carbon pyrophosphates. Twi 15 carbon farnesylpyrophosophate (FPP) molecules are the condensed by production of squalene (not show) to make the 30 carbon lanosterol, the first sterol in the pathway. This sterol is then used to make a variety of sterols, with cholesterol being the principle mammalian product, and ergosterol being that of yeast. The mevalonate pathway has many branches off the main pathway, delivering a wide variety of isroprene lipids for use in many corners of cell and organismal biology. One of these side branches included production of 20 carbon GGPP from 15 carbon FPP, discussed below. The key enzyme HMG-CoA reductase (HMGR) functions at the point indicated, catalyzing the reduction of the 6 carbon hydroxymethylglutaryl group on the CoA with 2NADPH to produce free mevalonic acid, as shown. To simplify the diagram, only the lipid structures are shown, while cofactors and non-lipids are absent.
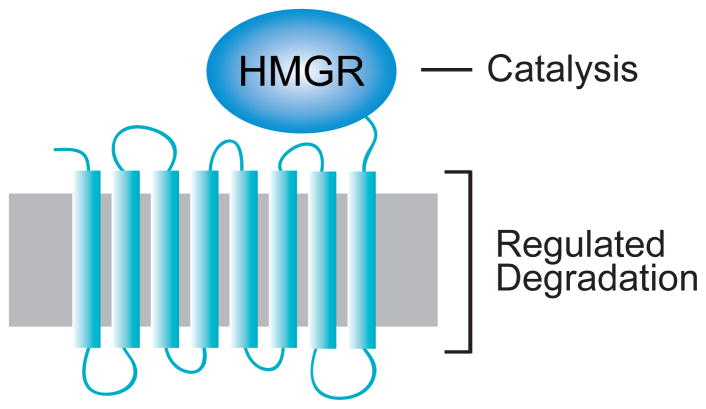
The integral ER membrane protein HMGR in both mammals and yeast consists of an 8-spanning N-terminal membrane anchor, embedded in the ER membrane (grey) as shown, attached by a linker to the highly conserved catalytic domain that performs the essential enzyme action. Regulated degradation requires the N-terminal transmembrane domain. This region has the conserved sterol-sensing domain (SSD) motif distributed between spans 2 to 5, as discussed in the text.
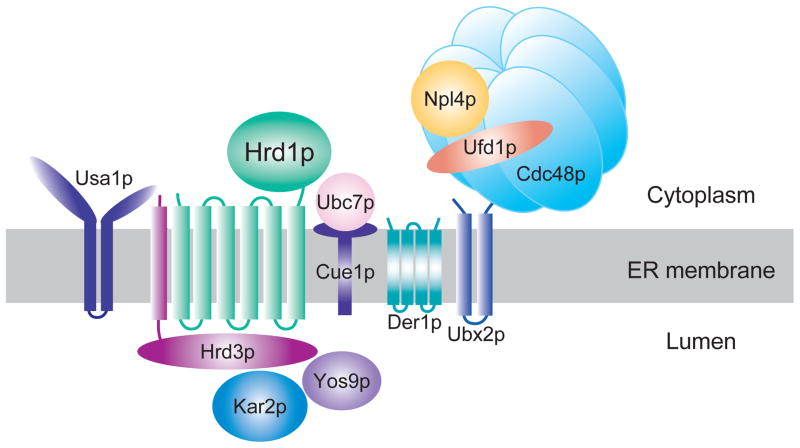
The HRD complex functions in ER-associated degradation (ERAD) to recognize and ubiquitinate misfolded and misassembled ER proteins. The Hrd1p E3 ubiquitin ligase works in a complex that is involved in all aspects of ERAD substrate recognition, ubiquitination, and movement to the cytosolic face where the proteasome resides. Hrd1p/Hrd3p are the E3 complex, and Ubc7p is the principle E2. The Cdc48 AAA-ATPase hexamer functions to remove substrates from the ER. Hrd3p, Kar2p, and Yos9p all function to recognized luminal ERAD substrates. The Usa1p protein regulates Hrd1p stability and activity along with allowing optimal Hrd1p activity. The HRD pathways has a large set of substrates that are recognized by a variety of criteria that hallmark poor folding.
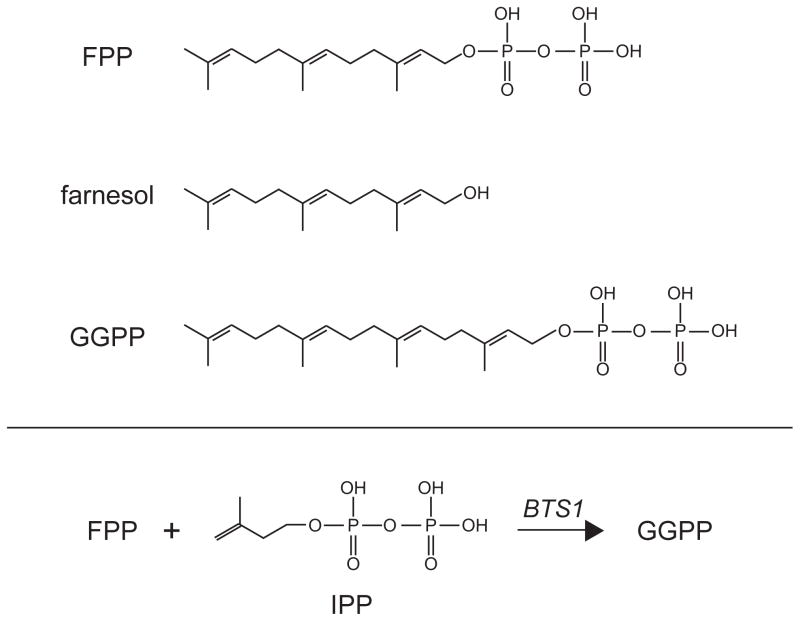
The structures of three isoprenes discussed in detail are shown, including 15 carbon farnesyl pyrophosphate (FPP), farnesol, and the 20 carbon geranylgeranyl pyrophosphate (GGPP). FPP levels in the cell are directly correlated with degradation rate of Hmg2p. Farnesol was employed in an the limited proteolysis assay to observe selective misfolding of the Hmg2p transmembrane domain. GGPP is featured in our most current studies on the signal that regulates Hmg2p degradation.
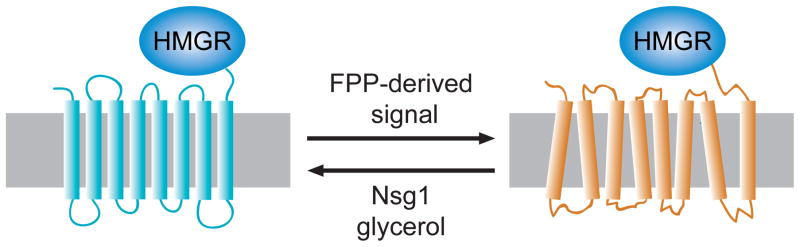
Regulated degradation of Hmg2p is posited to occur by a regulated change in structure to one that is more susceptible to HRD pathway degradation. Three ways of altering the stability of Hmg2p are indicated, with the direction of arrow showing the effect of each item. The FPP-derived signal causes more degradation, and it thought to altere the structure to one with more features of a misfolded protein. Conversely, Nsg1p or glycerol (a chemical chaperone) both effectively stabilize Hmg2p. Similarly, lowering the FPP-derived signal causes improved folding and thus, increased stability. The lower panel shows the metabolic reaction that generates GGPP from FPP by addition of 5 carbons from IPP, as shown. This reaction is catalyzed by the BTS1 gene product.
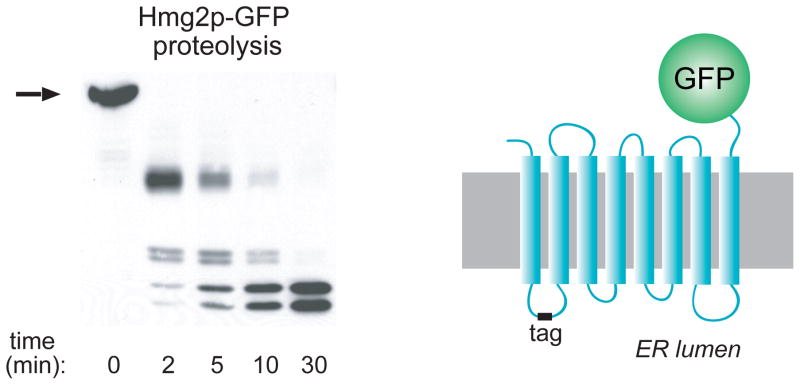
A myc epitope tag (“tag”) was added to the first lumenal loop of Hmg2p-GFP to create the normally regulated mycL-Hmg2p-GFP, depicted on the right. The tag is protected in the ER lumen, allowing detection of fragments generated during limited proteolysis of intact ER microsomes. A typical proteolysis experiment is shown in the left, in which microsomes are incubated with trypsin for the indicated times, and then subjected to SDS-PAGE and myc tag immunoblotting. All perturbations described in the text cause changes in the rate of proteolysis without any change in the order or pattern of fragment appearance. Stabilizing treatments such as chemical chaperones or preincubation with early sterol pathway inhibitors cause the proteolysis pattern to be generated more slowly. Conversely, treatments that cause more rapid degradation promote more rapid generation of the trypsinolysis pattern.
References
-
- Ciechanover A, Iwai K. IUBMB Life. 2004;56:193. - PubMed
-
- Richly H, Rape M, Braun S, Rumpf S, Hoege C, Jentsch S. Cell. 2005;120:73. - PubMed
-
- Elsasser S, Chandler-Militello D, Muller B, Hanna J, Finley D. J Biol Chem. 2004;279:26817. - PubMed
-
- Fang S, Jensen JP, Ludwig RL, Vousden KH, Weissman AM. J Biol Chem. 2000;275:8945. - PubMed
-
- Pines J. Trends Cell Biol. 2006;16:55. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
