

Innate and adaptive immunity for the pathobiology of Parkinson's disease
- PMID: 19243239
- PMCID: PMC2788126
- DOI: 10.1089/ars.2009.2460
Innate and adaptive immunity for the pathobiology of Parkinson's disease
Abstract
Innate and adaptive immunity affect the pathogenesis of Parkinson's disease (PD). In particular, activation of microglia influences degeneration of dopaminergic neurons. Cell-to-cell interactions and immune regulation critical for neuronal homeostasis also influence immune responses. The links between T cell immunity and nigrostriatal degeneration are supported by laboratory, animal model, and human pathologic investigations. Immune-associated biomarkers in spinal fluids and brain tissue of patients with idiopathic or familial forms of PD provide means to improve diagnosis and therapeutic monitoring. Relationships between oxidative stress, inflammation, and immune-mediated cell death pathways are examined in this review as they are linked to PD pathogenesis. Harnessing the immune system by drugs or by vaccination remain promising future therapeutic options.
Figures
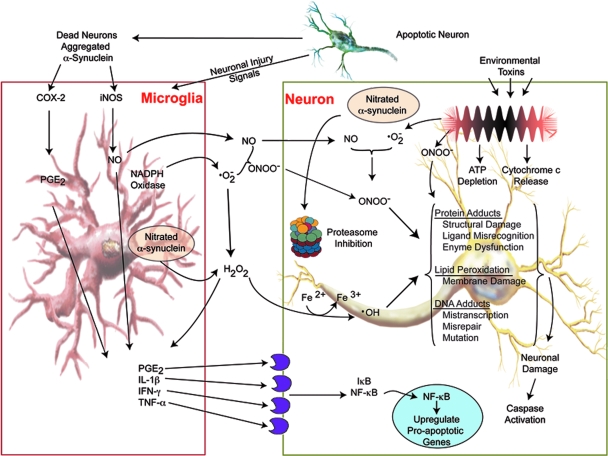
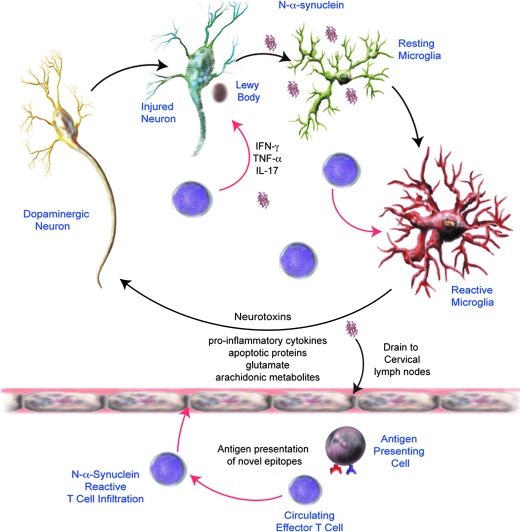
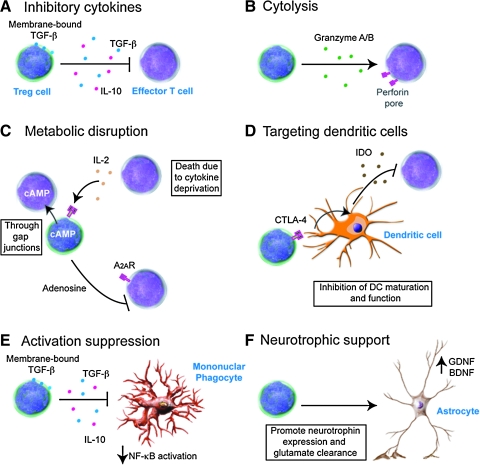
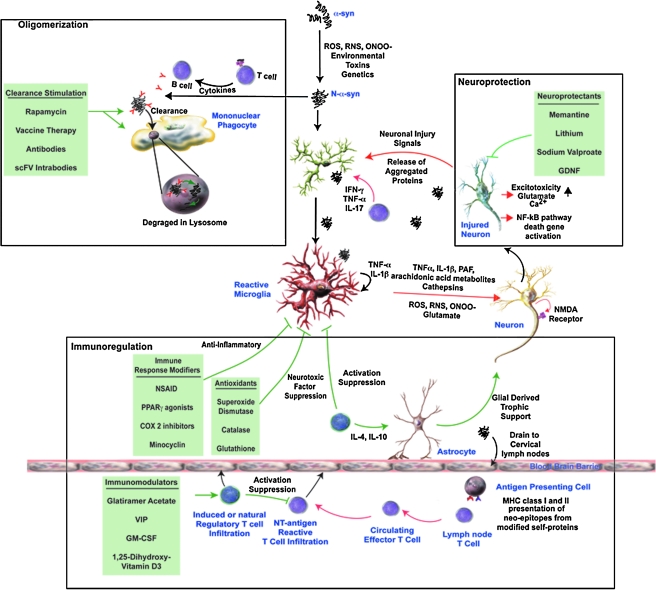
References
-
- abd-el-Basset E. Fedoroff S. Effect of bacterial wall lipopolysaccharide (LPS) on morphology, motility, and cytoskeletal organization of microglia in cultures. J Neurosci Res. 1995;41:222–237. - PubMed
-
- Aloisi F. Immune function of microglia. Glia. 2001;36:165–179. - PubMed
-
- Aloisi F. Penna G. Polazzi E. Minghetti L. Adorini L. CD40–CD154 interaction and IFN-gamma are required for IL-12 but not prostaglandin E2 secretion by microglia during antigen presentation to Th1 cells. J Immunol. 1999;162:1384–1391. - PubMed
-
- Amann R. Egger T. Schuligoi R. Heinemann A. Peskar BA. Sodium salicylate enhances the expression of cyclooxygenase-2 in endotoxin-stimulated human mononuclear cells. Eur J Pharmacol. 2001;433:129–134. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
