

Enhanced Rap1 activation and insulin secretagogue properties of an acetoxymethyl ester of an Epac-selective cyclic AMP analog in rat INS-1 cells: studies with 8-pCPT-2'-O-Me-cAMP-AM
- PMID: 19244230
- PMCID: PMC2667760
- DOI: 10.1074/jbc.M900166200
Enhanced Rap1 activation and insulin secretagogue properties of an acetoxymethyl ester of an Epac-selective cyclic AMP analog in rat INS-1 cells: studies with 8-pCPT-2'-O-Me-cAMP-AM
Abstract
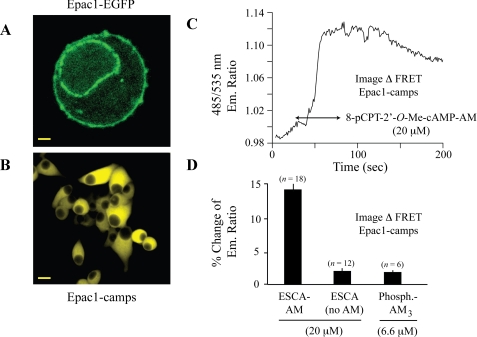
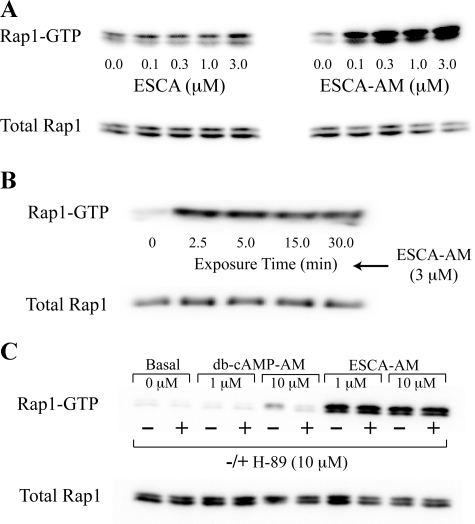
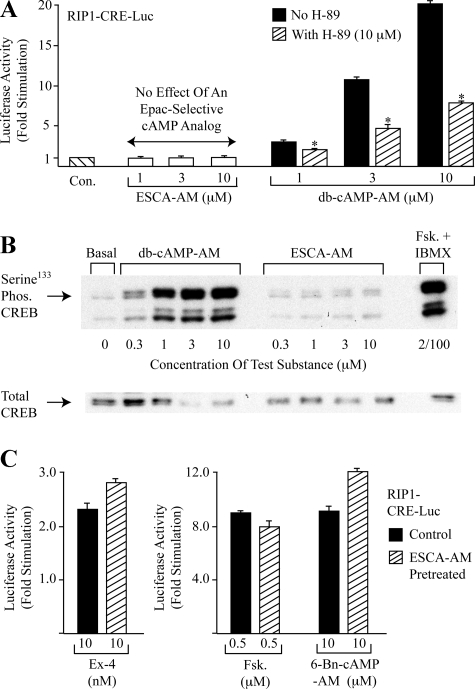
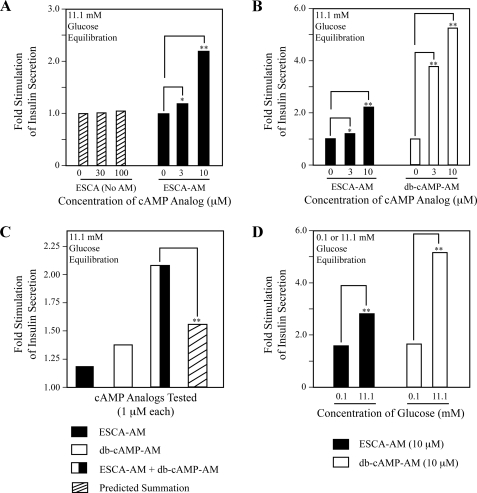
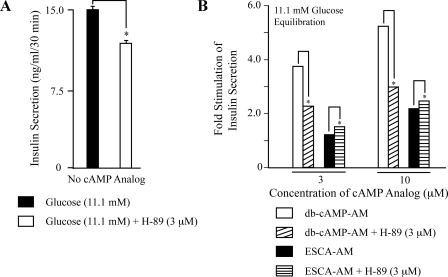
To ascertain the identities of cyclic nucleotide-binding proteins that mediate the insulin secretagogue action of cAMP, the possible contributions of the exchange protein directly activated by cAMP (Epac) and protein kinase A (PKA) were evaluated in a pancreatic beta cell line (rat INS-1 cells). Assays of Rap1 activation, CREB phosphorylation, and PKA-dependent gene expression were performed in combination with live cell imaging and high throughput screening of a fluorescence resonance energy transfer-based cAMP sensor (Epac1-camps) to validate the selectivity with which acetoxymethyl esters (AM-esters) of cAMP analogs preferentially activate Epac or PKA. Selective activation of Epac or PKA was achieved following exposure of INS-1 cells to 8-pCPT-2'-O-Me-cAMP-AM or Bt(2)cAMP-AM, respectively. Both cAMP analogs exerted dose-dependent and glucose metabolism-dependent actions to stimulate insulin secretion, and when each was co-administered with the other, a supra-additive effect was observed. Because 2.4-fold more insulin was secreted in response to a saturating concentration (10 microm) of Bt(2)cAMP-AM as compared with 8-pCPT-2'-O-Me-cAMP-AM, and because the action of Bt(2)cAMP-AM but not 8-pCPT-2'-O-Me-cAMP-AM was nearly abrogated by treatment with 3 microm of the PKA inhibitor H-89, it is concluded that for INS-1 cells, it is PKA that acts as the dominant cAMP-binding protein in support of insulin secretion. Unexpectedly, 10-100 microm of the non-AM-ester of 8-pCPT-2'-O-Me-cAMP failed to stimulate insulin secretion and was a weak activator of Rap1 in INS-1 cells. Moreover, 10 microm of the AM-ester of 8-pCPT-2'-O-Me-cAMP stimulated insulin secretion from mouse islets, whereas the non-AM-ester did not. Thus, the membrane permeability of 8-pCPT-2'-O-Me-cAMP in insulin-secreting cells is so low as to limit its biological activity. It is concluded that prior reports documenting the failure of 8-pCPT-2'-O-Me-cAMP to act in beta cells, or other cell types, need to be re-evaluated through the use of the AM-ester of this cAMP analog.
Figures

Similar articles
-
PKA-dependent potentiation of glucose-stimulated insulin secretion by Epac activator 8-pCPT-2'-O-Me-cAMP-AM in human islets of Langerhans.Am J Physiol Endocrinol Metab. 2010 Mar;298(3):E622-33. doi: 10.1152/ajpendo.00630.2009. Epub 2009 Dec 15. Am J Physiol Endocrinol Metab. 2010. PMID: 20009023 Free PMC article.
-
Phospholipase C-ε links Epac2 activation to the potentiation of glucose-stimulated insulin secretion from mouse islets of Langerhans.Islets. 2011 May-Jun;3(3):121-8. doi: 10.4161/isl.3.3.15507. Epub 2011 May 1. Islets. 2011. PMID: 21478675 Free PMC article.
-
Potentiation of sulfonylurea action by an EPAC-selective cAMP analog in INS-1 cells: comparison of tolbutamide and gliclazide and a potential role for EPAC activation of a 2-APB-sensitive Ca2+ influx.Mol Pharmacol. 2013 Jan;83(1):191-205. doi: 10.1124/mol.112.081943. Epub 2012 Oct 15. Mol Pharmacol. 2013. PMID: 23071106 Free PMC article.
-
Cooperation between cAMP signalling and sulfonylurea in insulin secretion.Diabetes Obes Metab. 2014 Sep;16 Suppl 1:118-25. doi: 10.1111/dom.12343. Diabetes Obes Metab. 2014. PMID: 25200305 Review.
-
Epac-selective cAMP analogs: new tools with which to evaluate the signal transduction properties of cAMP-regulated guanine nucleotide exchange factors.Cell Signal. 2008 Jan;20(1):10-20. doi: 10.1016/j.cellsig.2007.07.009. Epub 2007 Jul 25. Cell Signal. 2008. PMID: 17716863 Free PMC article. Review.
Cited by
-
Respiratory syncytial virus disrupts the airway epithelial barrier by decreasing cortactin and destabilizing F-actin.J Cell Sci. 2022 Aug 15;135(16):jcs259871. doi: 10.1242/jcs.259871. Epub 2022 Aug 16. J Cell Sci. 2022. PMID: 35848790 Free PMC article.
-
Epac2-dependent rap1 activation and the control of islet insulin secretion by glucagon-like peptide-1.Vitam Horm. 2010;84:279-302. doi: 10.1016/B978-0-12-381517-0.00010-2. Vitam Horm. 2010. PMID: 21094904 Free PMC article. Review.
-
Restoration of Glucose-Stimulated Cdc42-Pak1 Activation and Insulin Secretion by a Selective Epac Activator in Type 2 Diabetic Human Islets.Diabetes. 2018 Oct;67(10):1999-2011. doi: 10.2337/db17-1174. Epub 2018 Jul 9. Diabetes. 2018. PMID: 29986926 Free PMC article.
-
cAMP analogs and their metabolites enhance TREK-1 mRNA and K+ current expression in adrenocortical cells.Mol Pharmacol. 2010 Mar;77(3):469-82. doi: 10.1124/mol.109.061861. Epub 2009 Dec 22. Mol Pharmacol. 2010. PMID: 20028740 Free PMC article.
-
Glucose-dependent potentiation of mouse islet insulin secretion by Epac activator 8-pCPT-2'-O-Me-cAMP-AM.Islets. 2009 Nov-Dec;1(3):260-5. doi: 10.4161/isl.1.3.9645. Islets. 2009. PMID: 21099281 Free PMC article.
References
-
- de Rooij, J., Zwartkruis, F. J. T., Verheijen, M. H. G., Cool, R. H., Nijman, S. M. B., Wittinghofer, A., and Bos, J. L. (1998) Nature 396 474-477 - PubMed
-
- Kawasaki, H., Springett, G. M., Mochizuki, N., Toki, S., Nakaya, M., Matsuda, M., Housman, D. E., and Graybiel, A. M. (1998) Science 282 2275-2279 - PubMed
-
- Bos, J. L. (2006) Trends Biochem. Sci. 31 680-686 - PubMed
-
- Enserink, J. M., Christensen, A. E., de Rooij, J., Triest, M. V., Schwede, F., Genieser, H. G., Døskeland, S. O., Blank, J. L., and Bos, J. L. (2002) Nat. Cell Biol. 4 901-906 - PubMed
-
- Poppe, H., Rybalkin, S. D., Rehmann, H., Hinds, T. R., Tang, X. B., Christensen, A. E., Schwede, F., Genieser, H. G., Bos, J. L., Doskeland, S. O., Beavo, J. A., and Butt, E. (2008) Nat. Methods 5 277-288 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
