

Mice expressing an error-prone DNA polymerase in mitochondria display elevated replication pausing and chromosomal breakage at fragile sites of mitochondrial DNA
- PMID: 19244310
- PMCID: PMC2673436
- DOI: 10.1093/nar/gkp091
Mice expressing an error-prone DNA polymerase in mitochondria display elevated replication pausing and chromosomal breakage at fragile sites of mitochondrial DNA
Abstract
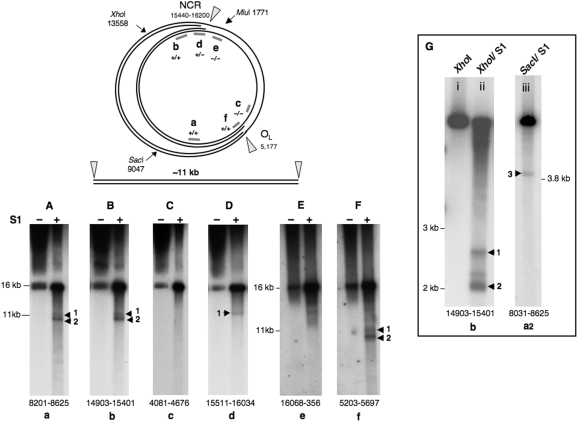
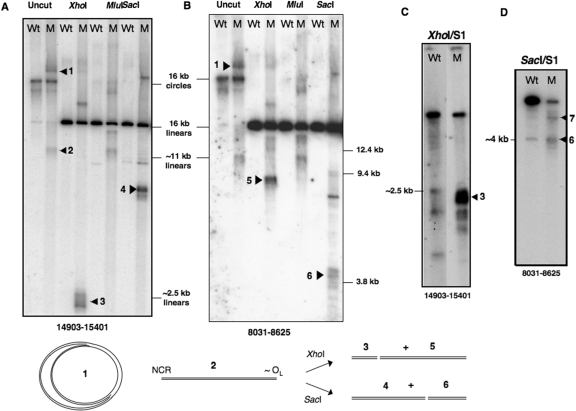
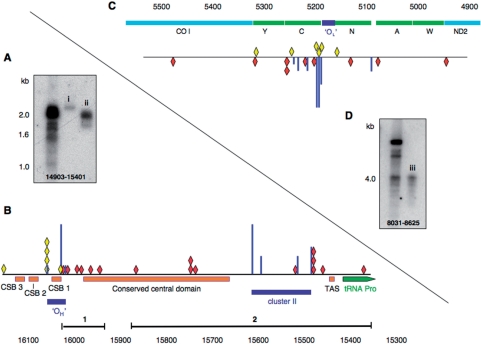
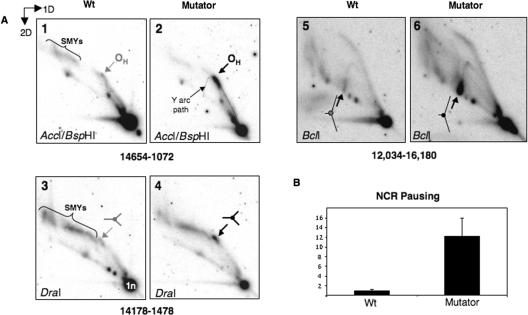
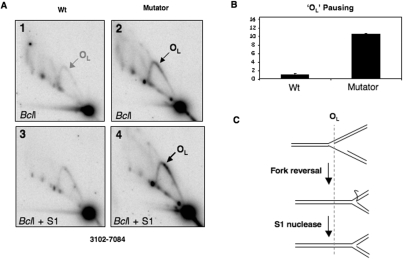
Expression of a proof-reading deficient form of mitochondrial DNA (mtDNA) polymerase gamma, POLG, causes early death accompanied by features of premature ageing in mouse. However, the mechanism of cellular senescence remains unresolved. In addition to high levels of point mutations of mtDNA, the POLG mutator mouse harbours linear mtDNAs. Using one- and two-dimensional agarose gel electrophoresis, we show that the linear mtDNAs derive from replication intermediates and are indicative of replication pausing and chromosomal breakage at the accompanying fragile sites. Replication fork arrest is not random but occurs at specific sites close to two cis-elements known as O(H) and O(L). Pausing at these sites may be enhanced in the case of exonuclease-deficient POLG owing to delayed resumption of DNA replication, or replisome instability. In either case, the mtDNA replication cycle is perturbed and this might explain the progeroid features of the POLG mutator mouse.
Figures
Similar articles
-
The exonuclease activity of DNA polymerase γ is required for ligation during mitochondrial DNA replication.Nat Commun. 2015 Jun 22;6:7303. doi: 10.1038/ncomms8303. Nat Commun. 2015. PMID: 26095671 Free PMC article.
-
The mtDNA mutation spectrum of the progeroid Polg mutator mouse includes abundant control region multimers.Cell Metab. 2010 Dec 1;12(6):675-82. doi: 10.1016/j.cmet.2010.11.012. Cell Metab. 2010. PMID: 21109200 Free PMC article.
-
In vivo functional analysis of the human mitochondrial DNA polymerase POLG expressed in cultured human cells.J Biol Chem. 2000 Aug 11;275(32):24818-28. doi: 10.1074/jbc.M000559200. J Biol Chem. 2000. PMID: 10827171
-
The mitochondrial DNA polymerase in health and disease.Subcell Biochem. 2010;50:211-22. doi: 10.1007/978-90-481-3471-7_11. Subcell Biochem. 2010. PMID: 20012584 Free PMC article. Review.
-
Consequences of compromised mitochondrial genome integrity.DNA Repair (Amst). 2020 Sep;93:102916. doi: 10.1016/j.dnarep.2020.102916. DNA Repair (Amst). 2020. PMID: 33087282 Free PMC article. Review.
Cited by
-
Is There Still Any Role for Oxidative Stress in Mitochondrial DNA-Dependent Aging?Genes (Basel). 2018 Mar 21;9(4):175. doi: 10.3390/genes9040175. Genes (Basel). 2018. PMID: 29561808 Free PMC article. Review.
-
Ultra-deep sequencing of mouse mitochondrial DNA: mutational patterns and their origins.PLoS Genet. 2011 Mar;7(3):e1002028. doi: 10.1371/journal.pgen.1002028. Epub 2011 Mar 24. PLoS Genet. 2011. PMID: 21455489 Free PMC article.
-
Association of genetic variations in the mitochondrial DNA control region with presbycusis.Clin Interv Aging. 2017 Mar 3;12:459-465. doi: 10.2147/CIA.S123278. eCollection 2017. Clin Interv Aging. 2017. PMID: 28424544 Free PMC article.
-
Mitochondrial DNA alterations and reduced mitochondrial function in aging.Mech Ageing Dev. 2010 Jul-Aug;131(7-8):451-62. doi: 10.1016/j.mad.2010.03.007. Epub 2010 Mar 20. Mech Ageing Dev. 2010. PMID: 20307565 Free PMC article. Review.
-
Tissue-specific responses to TFAM and mtDNA copy number manipulation in prematurely ageing mice.Elife. 2025 Jun 30;14:RP104461. doi: 10.7554/eLife.104461. Elife. 2025. PMID: 40587199 Free PMC article.
References
-
- Li Y, Huang TT, Carlson EJ, Melov S, Ursell PC, Olson JL, Noble LJ, Yoshimura MP, Berger C, Chan PH, et al. Dilated cardiomyopathy and neonatal lethality in mutant mice lacking manganese superoxide dismutase. Nat. Genet. 1995;11:376–381. - PubMed
-
- Reaume AG, Elliott JL, Hoffman EK, Kowall NW, Ferrante RJ, Siwek DF, Wilcox HM, Flood DG, Beal MF, Brown RH, Jr., et al. Motor neurons in Cu/Zn superoxide dismutase-deficient mice develop normally but exhibit enhanced cell death after axonal injury. Nat. Genet. 1996;13:43–47. - PubMed
-
- Schriner SE, Linford NJ, Martin GM, Treuting P, Ogburn CE, Emond M, Coskun PE, Ladiges W, Wolf N, Van Remmen H, et al. Extension of murine life span by overexpression of catalase targeted to mitochondria. Science. 2005;308:1909–1911. - PubMed
-
- Jacobs HT. The mitochondrial theory of aging: dead or alive? Aging Cell. 2003;2:11–17. - PubMed
-
- Bender A, Krishnan KJ, Morris CM, Taylor GA, Reeve AK, Perry RH, Jaros E, Hersheson JS, Betts J, Klopstock T, et al. High levels of mitochondrial DNA deletions in substantia nigra neurons in aging and Parkinson disease. Nat. Genet. 2006;38:515–517. - PubMed
