

NF-kappaB serves as a cellular sensor of Kaposi's sarcoma-associated herpesvirus latency and negatively regulates K-Rta by antagonizing the RBP-Jkappa coactivator
- PMID: 19244329
- PMCID: PMC2668470
- DOI: 10.1128/JVI.01999-08
NF-kappaB serves as a cellular sensor of Kaposi's sarcoma-associated herpesvirus latency and negatively regulates K-Rta by antagonizing the RBP-Jkappa coactivator
Abstract
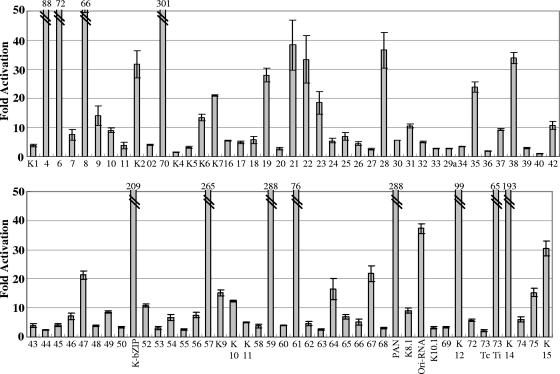
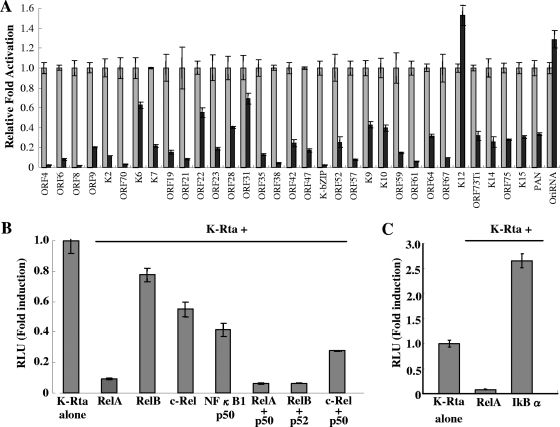
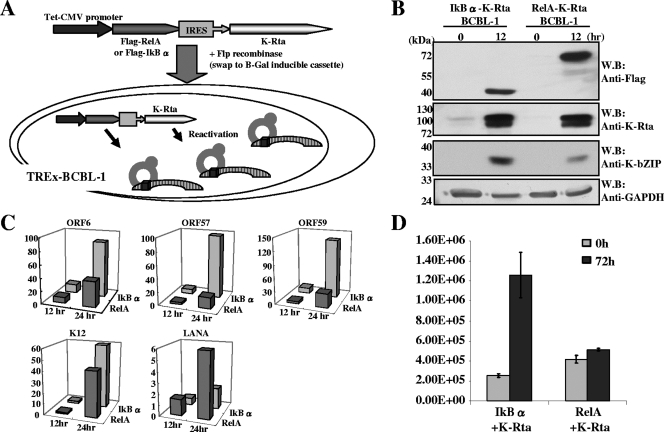
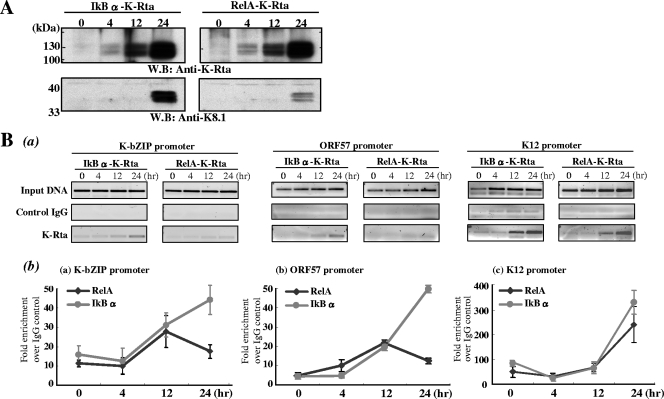
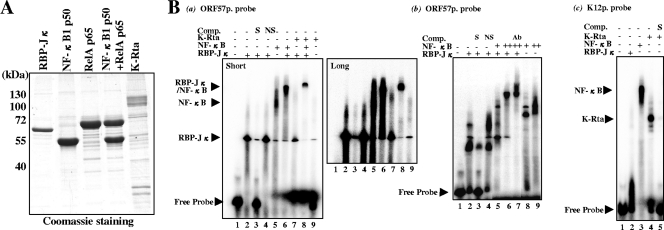
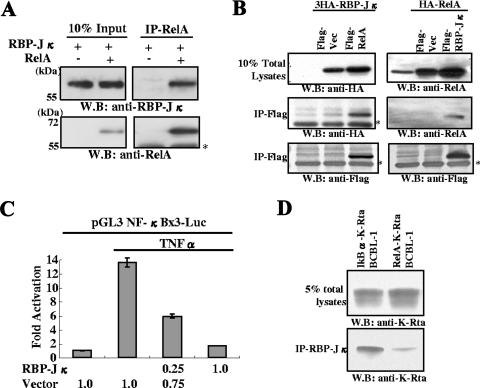
Successful viral replication is dependent on a conducive cellular environment; thus, viruses must be sensitive to the state of their host cells. We examined the idea that an interplay between viral and cellular regulatory factors determines the switch from Kaposi's sarcoma-associated herpesvirus (KSHV) latency to lytic replication. The immediate-early gene product K-Rta is the first viral protein expressed and an essential factor in reactivation; accordingly, this viral protein is in a key position to serve as a viral sensor of cellular physiology. Our approach aimed to define a host transcription factor, i.e., host sensor, which modulates K-Rta activity on viral promoters. To this end, we developed a panel of reporter plasmids containing all 83 putative viral promoters for a comprehensive survey of the response to both K-Rta and cellular transcription factors. Interestingly, members of the NF-kappaB family were shown to be strong negative regulators of K-Rta transactivation for all but two viral promoters (Ori-RNA and K12). Recruitment of K-Rta to the ORF57 and K-bZIP promoters, but not the K12 promoter, was significantly impaired when NF-kappaB expression was induced. Many K-Rta-responsive promoters modulated by NF-kappaB contain the sequence of the RBP-Jkappa binding site, a major coactivator which anchors K-Rta to target promoters via consensus motifs which overlap with that of NF-kappaB. Gel shift assays demonstrated that NF-kappaB inhibits the binding of RBP-Jkappa and forms a complex with RBP-Jkappa. Our results support a model in which a balance between K-Rta/RBP-Jkappa and NF-kappaB activities determines KSHV reactivation. An important feature of this model is that the interplay between RBP-Jkappa and NF-kappaB on viral promoters controls viral gene expression mediated by K-Rta.
Figures
Similar articles
-
Genome-Wide Identification of Direct RTA Targets Reveals Key Host Factors for Kaposi's Sarcoma-Associated Herpesvirus Lytic Reactivation.J Virol. 2019 Feb 19;93(5):e01978-18. doi: 10.1128/JVI.01978-18. Print 2019 Mar 1. J Virol. 2019. PMID: 30541837 Free PMC article.
-
Kaposi's sarcoma-associated herpesvirus reactivation is regulated by interaction of latency-associated nuclear antigen with recombination signal sequence-binding protein Jkappa, the major downstream effector of the Notch signaling pathway.J Virol. 2005 Mar;79(6):3468-78. doi: 10.1128/JVI.79.6.3468-3478.2005. J Virol. 2005. PMID: 15731241 Free PMC article.
-
Direct interactions of Kaposi's sarcoma-associated herpesvirus/human herpesvirus 8 ORF50/Rta protein with the cellular protein octamer-1 and DNA are critical for specifying transactivation of a delayed-early promoter and stimulating viral reactivation.J Virol. 2007 Aug;81(16):8451-67. doi: 10.1128/JVI.00265-07. Epub 2007 May 30. J Virol. 2007. PMID: 17537858 Free PMC article.
-
KSHV reactivation and novel implications of protein isomerization on lytic switch control.Viruses. 2015 Jan 12;7(1):72-109. doi: 10.3390/v7010072. Viruses. 2015. PMID: 25588053 Free PMC article. Review.
-
KSHV miRNAs decrease expression of lytic genes in latently infected PEL and endothelial cells by targeting host transcription factors.Viruses. 2014 Oct 23;6(10):4005-23. doi: 10.3390/v6104005. Viruses. 2014. PMID: 25341664 Free PMC article. Review.
Cited by
-
Kaposi's Sarcoma-Associated Herpesvirus Hijacks RNA Polymerase II To Create a Viral Transcriptional Factory.J Virol. 2017 May 12;91(11):e02491-16. doi: 10.1128/JVI.02491-16. Print 2017 Jun 1. J Virol. 2017. PMID: 28331082 Free PMC article.
-
Regulatory network mediated by RBP-J/NFATc1-miR182 controls inflammatory bone resorption.FASEB J. 2020 Feb;34(2):2392-2407. doi: 10.1096/fj.201902227R. Epub 2019 Dec 23. FASEB J. 2020. PMID: 31908034 Free PMC article.
-
KSHV transactivator-derived small peptide traps coactivators to attenuate MYC and inhibits leukemia and lymphoma cell growth.Commun Biol. 2021 Dec 2;4(1):1330. doi: 10.1038/s42003-021-02853-0. Commun Biol. 2021. PMID: 34857874 Free PMC article.
-
miR-K12-7-5p encoded by Kaposi's sarcoma-associated herpesvirus stabilizes the latent state by targeting viral ORF50/RTA.PLoS One. 2011 Jan 20;6(1):e16224. doi: 10.1371/journal.pone.0016224. PLoS One. 2011. PMID: 21283761 Free PMC article.
-
KSHV vIL-6 enhances inflammatory responses by epigenetic reprogramming.PLoS Pathog. 2023 Nov 7;19(11):e1011771. doi: 10.1371/journal.ppat.1011771. eCollection 2023 Nov. PLoS Pathog. 2023. PMID: 37934757 Free PMC article.
References
-
- Carroll, K. D., F. Khadim, S. Spadavecchia, D. Palmeri, and D. M. Lukac. 2007. Direct interactions of Kaposi's sarcoma-associated herpesvirus/human herpesvirus 8 ORF50/Rta protein with the cellular protein octamer-1 and DNA are critical for specifying transactivation of a delayed-early promoter and stimulating viral reactivation. J. Virol. 818451-8467. - PMC - PubMed
-
- Caselli, E., S. Fiorentini, C. Amici, D. Di Luca, A. Caruso, and M. G. Santoro. 2007. Human herpesvirus 8 acute infection of endothelial cells induces monocyte chemoattractant protein 1-dependent capillary-like structure formation: role of the IKK/NF-κB pathway. Blood 1092718-2726. - PubMed
