

Entry of herpes simplex virus 1 and other alphaherpesviruses via the paired immunoglobulin-like type 2 receptor alpha
- PMID: 19244335
- PMCID: PMC2668467
- DOI: 10.1128/JVI.02601-08
Entry of herpes simplex virus 1 and other alphaherpesviruses via the paired immunoglobulin-like type 2 receptor alpha
Abstract
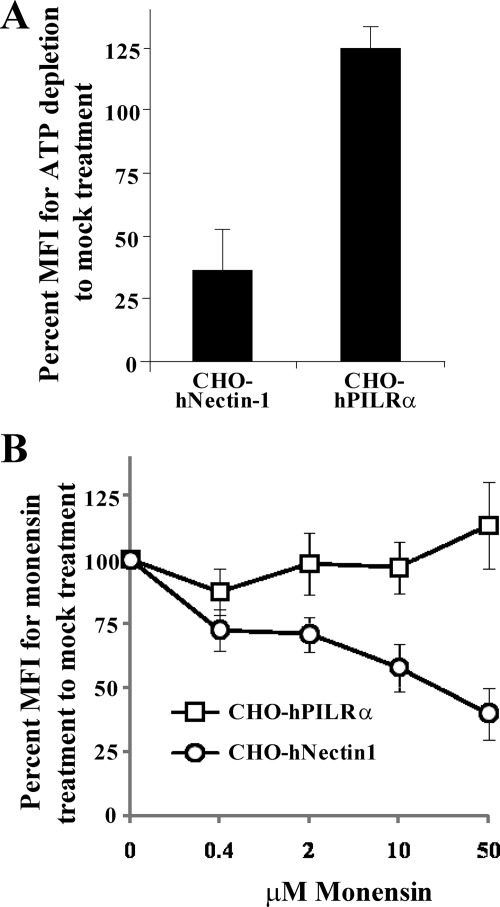
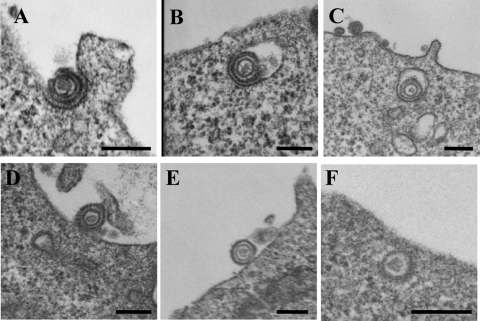
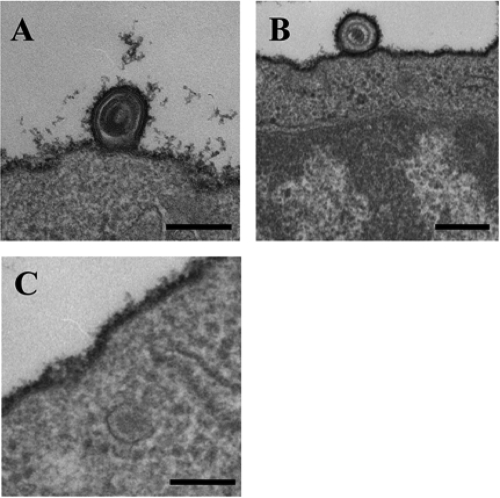
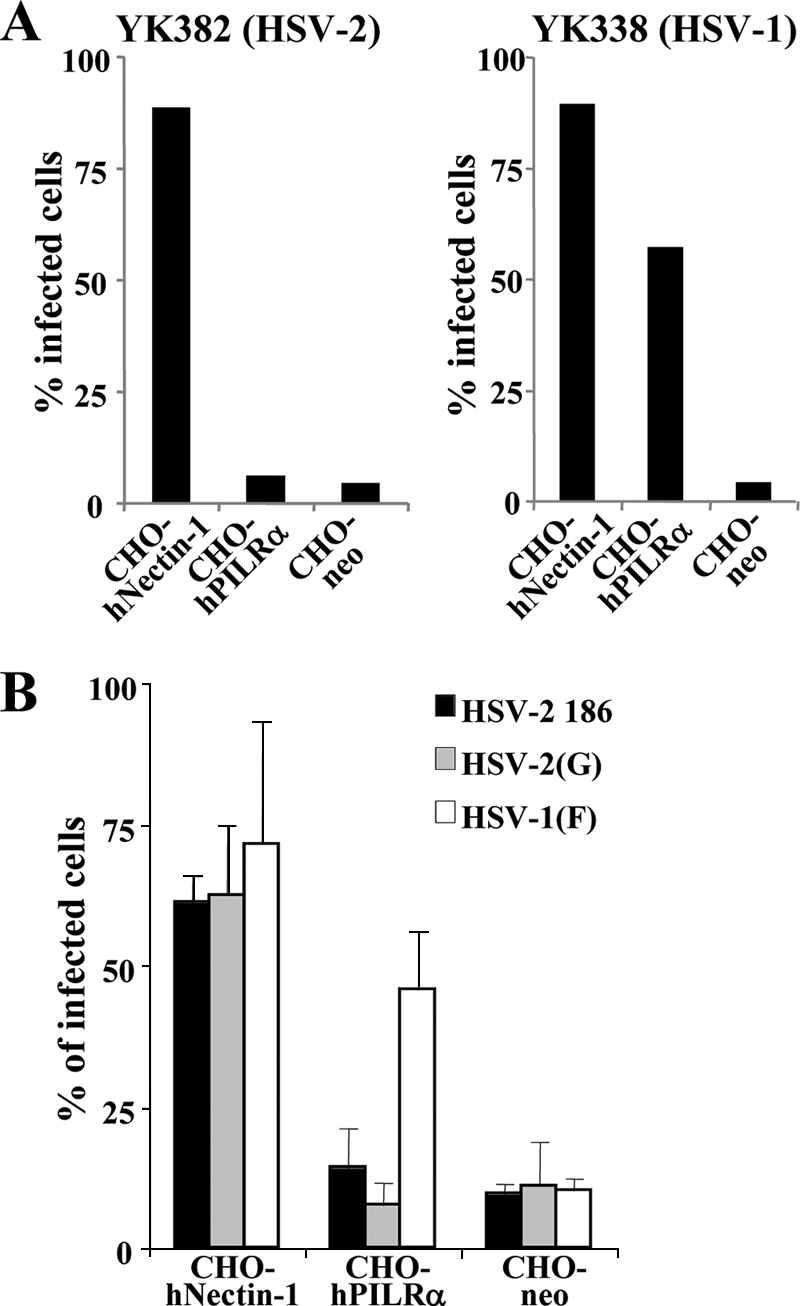
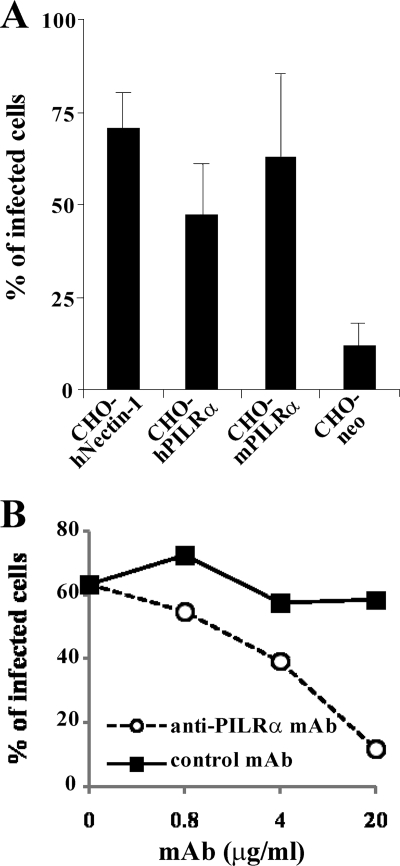
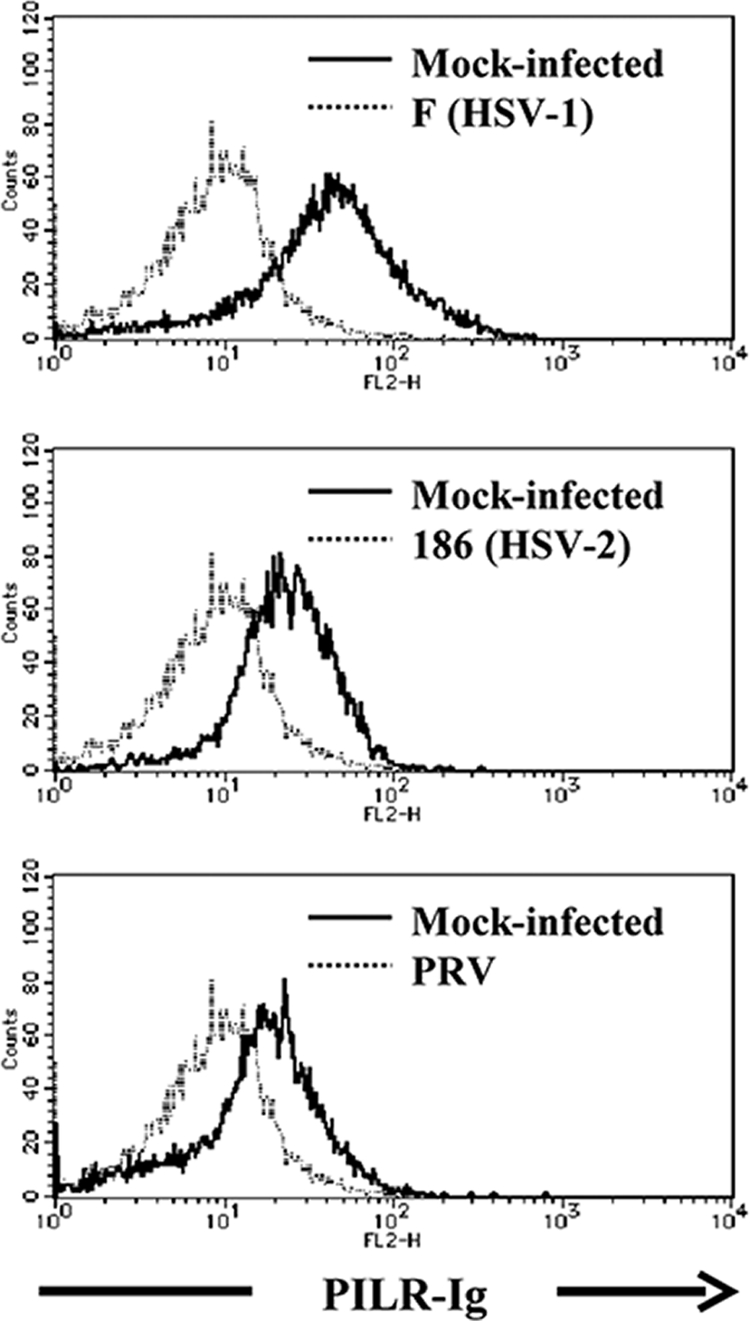
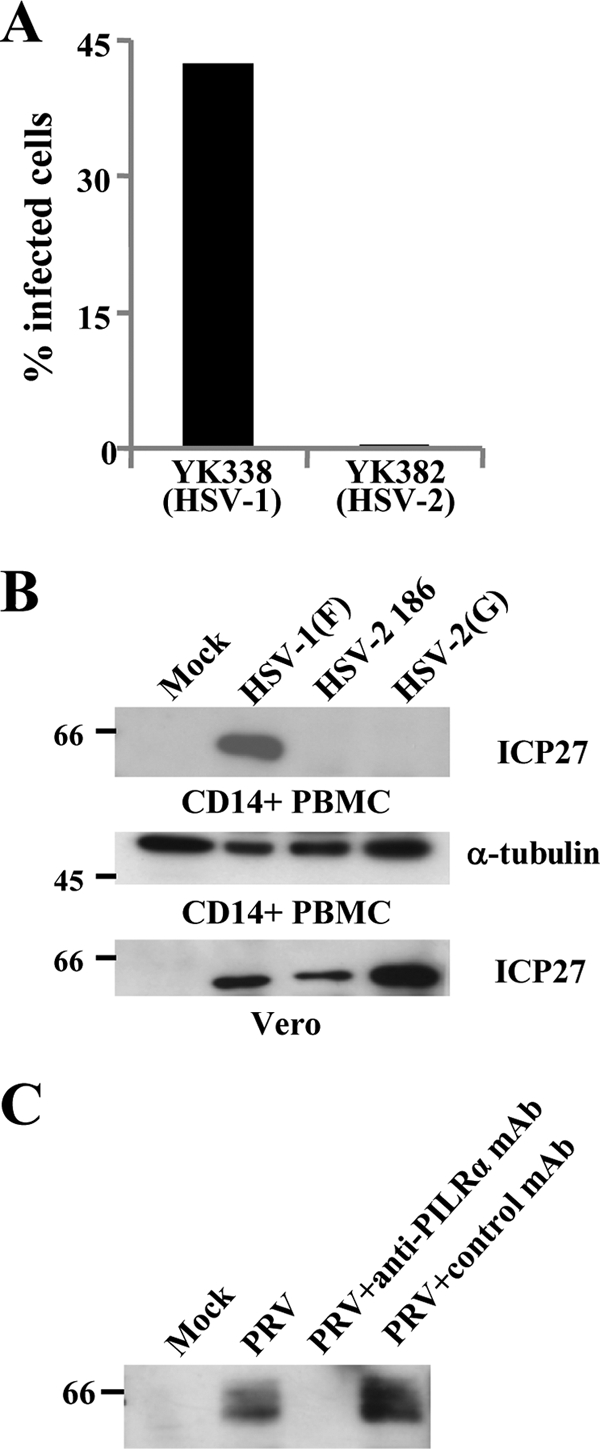
Herpes simplex virus 1 (HSV-1) enters cells either via fusion of the virion envelope and host cell plasma membrane or via endocytosis, depending on the cell type. In the study reported here, we investigated a viral entry pathway dependent on the paired immunoglobulin-like type 2 receptor alpha (PILRalpha), a recently identified entry coreceptor for HSV-1 that associates with viral envelope glycoprotein B (gB). Experiments using inhibitors of endocytic pathways and ultrastructural analyses of Chinese hamster ovary (CHO) cells transduced with PILRalpha showed that HSV-1 entry into these cells was via virus-cell fusion at the cell surface. Together with earlier observations that HSV-1 uptake into normal CHO cells and those transduced with a receptor for HSV-1 envelope gD is mediated by endocytosis, these results indicated that expression of PILRalpha produced an alternative HSV-1 entry pathway in CHO cells. We also showed that human and murine PILRalpha were able to mediate entry of pseudorabies virus, a porcine alphaherpesvirus, but not of HSV-2. These results indicated that viral entry via PILRalpha appears to be conserved but that there is a PILRalpha preference among alphaherpesviruses.
Figures
References
-
- Ahmad, R., J. Ennaciri, P. Cordeiro, S. El Bassam, and J. Menezes. 2007. Herpes simplex virus-1 up-regulates IL-15 gene expression in monocytic cells through the activation of protein tyrosine kinase and PKC zeta/lambda signaling pathways. J. Mol. Biol. 36725-35. - PubMed
-
- Bosnjak, L., M. Miranda-Saksena, D. M. Koelle, R. A. Boadle, C. A. Jones, and A. L. Cunningham. 2005. Herpes simplex virus infection of human dendritic cells induces apoptosis and allows cross-presentation via uninfected dendritic cells. J. Immunol. 1742220-2227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
