

Differential 3' splice site recognition of SMN1 and SMN2 transcripts by U2AF and U2 snRNP
- PMID: 19244360
- PMCID: PMC2661831
- DOI: 10.1261/rna.1273209
Differential 3' splice site recognition of SMN1 and SMN2 transcripts by U2AF and U2 snRNP
Abstract
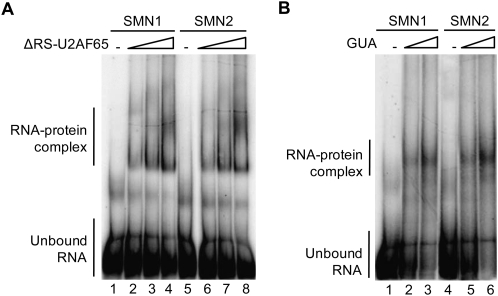
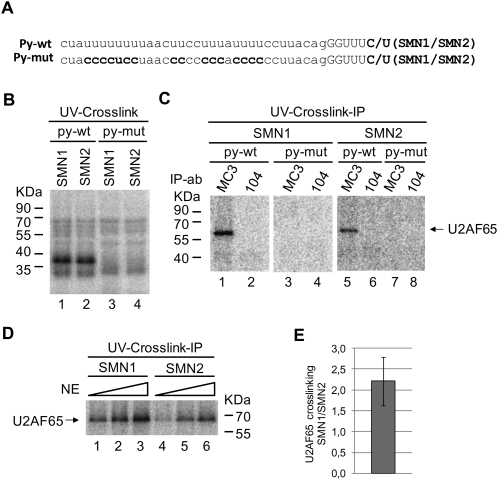
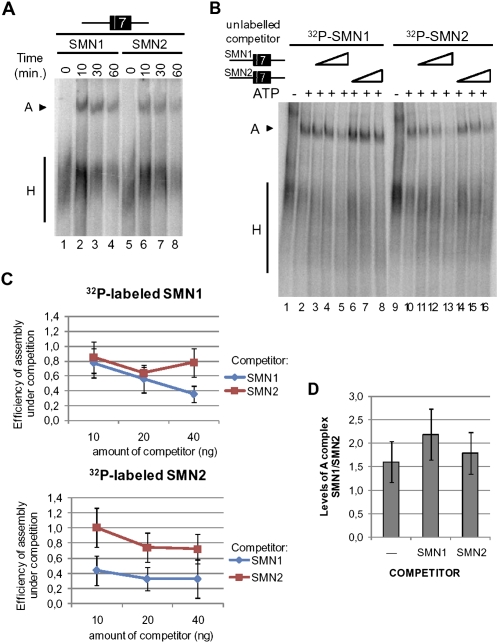
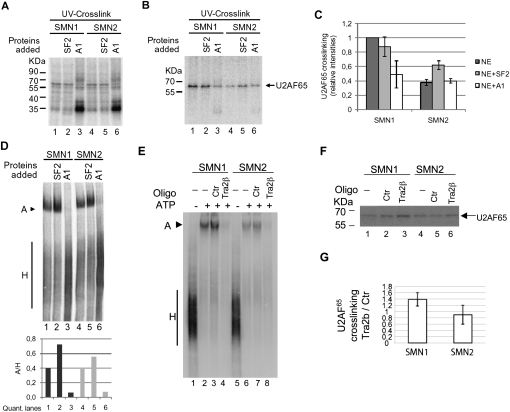
Spinal Muscular atrophy is a prevalent genetic disease caused by mutation of the SMN1 gene, which encodes the SMN protein involved in assembly of small nuclear ribonucleoprotein (snRNP) complexes. A paralog of the gene, SMN2, cannot provide adequate levels of functional SMN because exon 7 is skipped in a significant fraction of the mature transcripts. A C to T transition located at position 6 of exon 7 is critical for the difference in exon skipping between SMN1 and SMN2. Here we report that this nucleotide difference results in increased ultraviolet light-mediated crosslinking of the splicing factor U2AF(65) with the 3' splice site of SMN1 intron 6 in HeLa nuclear extract. U2 snRNP association, analyzed by native gel electrophoresis, is also more efficient on SMN1 than on SMN2, particularly under conditions of competition, suggesting more effective use of limiting factors. Two trans-acting factors implicated in SMN regulation, SF2/ASF and hnRNP A1, promote and repress, respectively, U2 snRNP recruitment to both RNAs. Interestingly, depending on the transcript and the regulatory factor, the effects on U2 binding not always correlate with changes in U2AF(65) crosslinking. Furthermore, blocking recognition of a Tra2-beta1-dependent splicing enhancer located in exon 7 inhibits U2 snRNP recruitment without affecting U2AF(65) crosslinking. Collectively, the results suggest that both U2AF binding and other steps of U2 snRNP recruitment can be control points in SMN splicing regulation.
Figures
References
-
- Bonnal S., Martínez C., Förch P., Bachi A., Wilm M., Valcárcel J. RBM5/Luca-15/H37 regulates Fas alternative splice site pairing after exon definition. Mol. Cell. 2008;32:81–95. - PubMed
-
- Brzustowicz L.M., Lehner T., Castilla L.H., Penchaszadeh G.K., Wilhelmsen K.C., Daniels R., Davies K.E., Leppert M., Ziter F., Wood D., et al. Genetic mapping of chronic childhood-onset spinal muscular atrophy to chromosome 5q11.2–13.3. Nature. 1990;344:540–541. - PubMed
-
- Cartegni L., Krainer A.R. Disruption of an SF2/ASF-dependent exonic splicing enhancer in SMN2 causes spinal muscular atrophy in the absence of SMN1. Nat. Genet. 2002;30:377–384. - PubMed
-
- Cartegni L., Krainer A.R. Correction of disease-associated exon skipping by synthetic exon-specific activators. Nat. Struct. Biol. 2003;10:120–125. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources