

Substrate-triggered formation and remarkable stability of the C-H bond-cleaving chloroferryl intermediate in the aliphatic halogenase, SyrB2
- PMID: 19245217
- PMCID: PMC2684568
- DOI: 10.1021/bi900109z
Substrate-triggered formation and remarkable stability of the C-H bond-cleaving chloroferryl intermediate in the aliphatic halogenase, SyrB2
Abstract
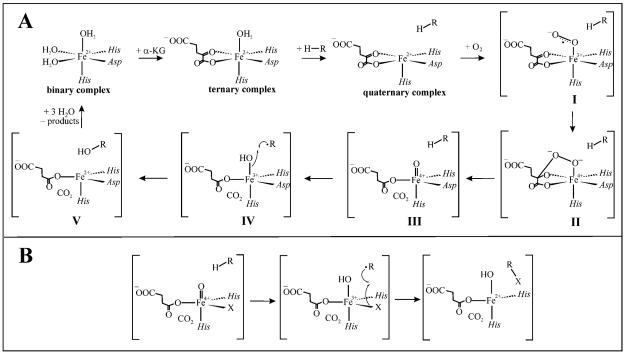
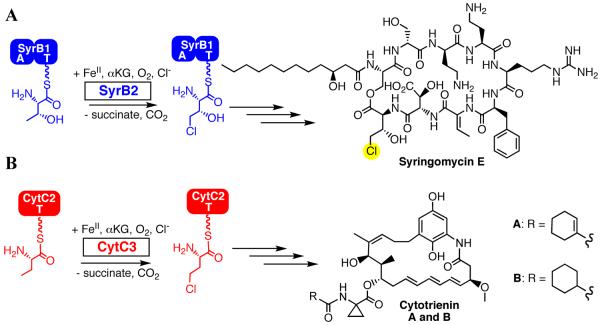
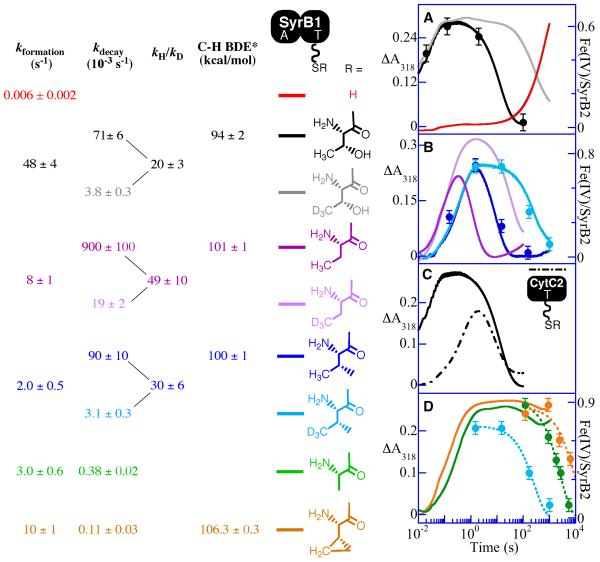
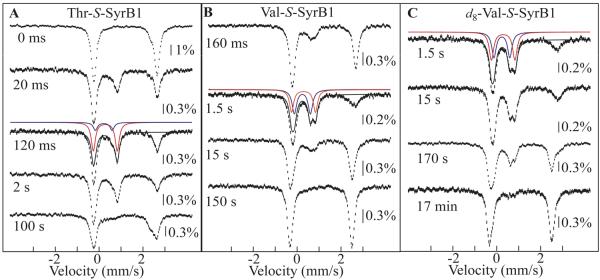
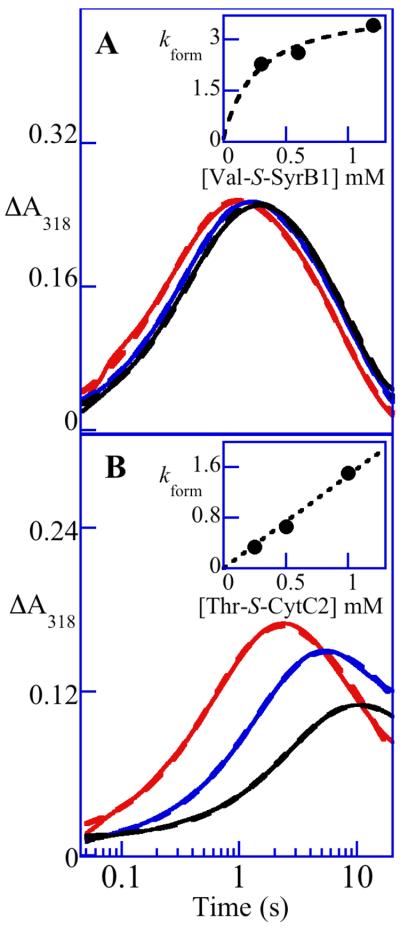
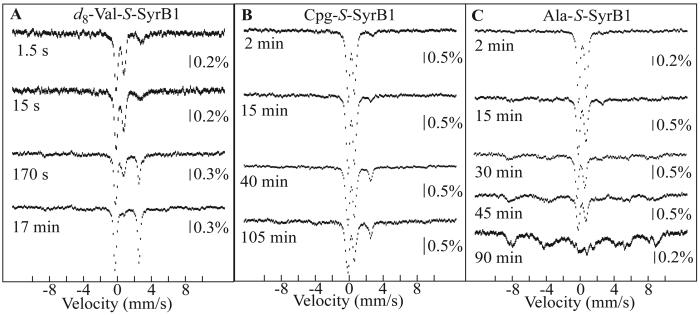
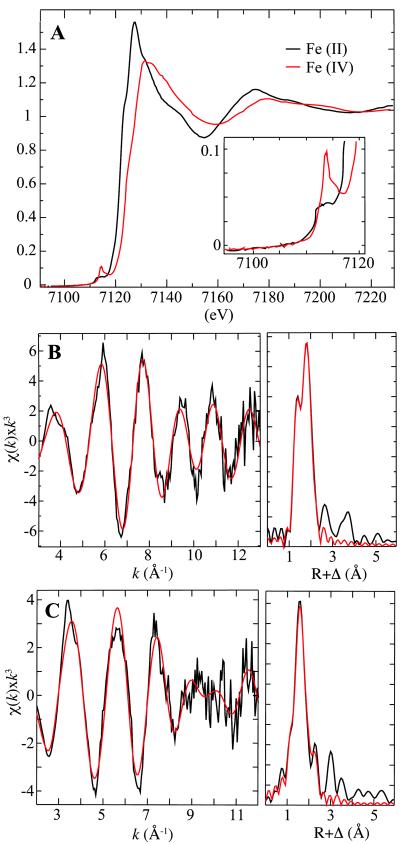
Aliphatic halogenases activate O(2), cleave alpha-ketoglutarate (alphaKG) to CO(2) and succinate, and form haloferryl [X-Fe(IV)O; X = Cl or Br] complexes that cleave aliphatic C-H bonds to install halogens during the biosynthesis of natural products by non-ribosomal peptide synthetases (NRPSs). For the related alphaKG-dependent dioxygenases, it has been shown that reaction of the Fe(II) cofactor with O(2) to form the C-H bond-cleaving ferryl complex is "triggered" by binding of the target substrate. In this study, we have tested for and defined structural determinants of substrate triggering (ST) in the halogenase, SyrB2, from the syringomycin E biosynthetic NRPS of Pseudomonas syringae B301D. As for other halogenases, the substrate of SyrB2 is complex, consisting of l-Thr tethered via a thioester linkage to a covalently bound phosphopantetheine (PPant) cofactor of a carrier protein, SyrB1. Without an appended amino acid, SyrB1 does not trigger formation of the chloroferryl intermediate state in SyrB2, even in the presence of free l-Thr or its analogues, but SyrB1 charged either by l-Thr (l-Thr-S-SyrB1) or by any of several non-native amino acids does trigger the reaction by as much as 8000-fold (for the native substrate). Triggering efficacy is sensitive to the structures of both the amino acid and the carrier protein, being diminished by 5-24-fold when the native l-Thr is replaced with another amino acid and by approximately 40-fold when SyrB1 is replaced with the heterologous carrier protein, CytC2. The directing effect of the carrier protein and consequent tolerance for profound modifications to the target amino acid allow the chloroferryl state to be formed in the presence of substrates that perturb the ratio of its two putative coordination isomers, lack the target C-H bond (l-Ala-S-SyrB1), or contain a C-H bond of enhanced strength (l-cyclopropylglycyl-S-SyrB1). For the latter two cases, the SyrB2 chloroferryl state so formed exhibits unprecedented stability (t(1/2) = 30-110 min at 0 degree C), can be trapped at high concentration and purity by manual freezing without a cryosolvent, and represents an ideal target for structural characterization. As initial steps toward this goal, extended X-ray absorption fine structure (EXAFS) spectroscopy has been used to determine the Fe-O and Fe-Cl distances and density functional theory (DFT) calculations have been used to confirm that the measured distances are consistent with the anticipated structure of the intermediate.
Figures
References
-
- Hausinger RP. Fe(II)/alpha-ketoglutarate-dependent hydroxylases and related enzymes. Crit. Rev. Biochem. Mol. Biol. 2004;39:21–68. - PubMed
-
- Solomon EI, Brunold TC, Davis MI, Kemsley JN, Lee S-K, Lehnert N, Neese F, Skulan AJ, Yang Y-S, Zhou J. Geometric and electronic structure/function correlations in non-heme iron enzymes. Chem. Rev. 2000;100:235–349. - PubMed
-
- Costas M, Mehn MP, Jensen MP, Que L., Jr. Dioxygen activation at mononuclear nonheme iron active sites: Enzymes, models, and intermediates. Chem. Rev. 2004;104:939–986. - PubMed
-
- Que L., Jr. One motif - many different reactions. Nature Struct. Biol. 2000;7:182–184. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
