

Telomeres do the (un)twist: helicase actions at chromosome termini
- PMID: 19245831
- PMCID: PMC2670356
- DOI: 10.1016/j.bbadis.2009.02.008
Telomeres do the (un)twist: helicase actions at chromosome termini
Abstract
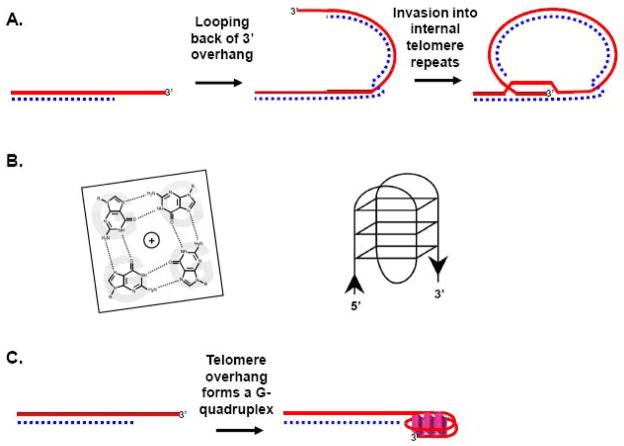
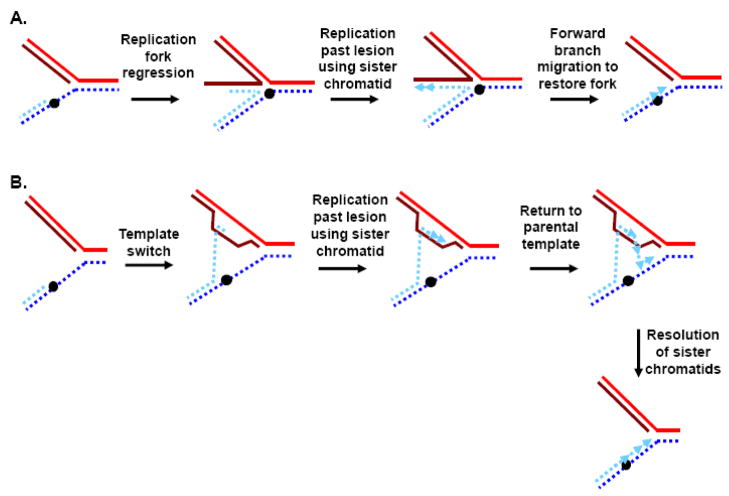
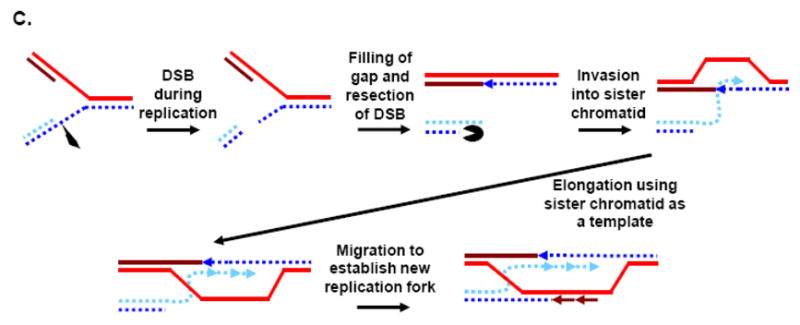
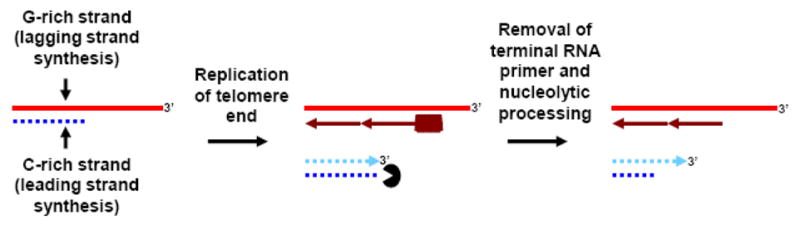
Telomeres play critical roles in protecting genome stability, and their dysfunction contributes to cancer and age-related degenerative diseases. The precise architecture of telomeres, including their single-stranded 3' overhangs, bound proteins, and ability to form unusual secondary structures such as t-loops, is central to their function and thus requires careful processing by diverse factors. Furthermore, telomeres provide unique challenges to the DNA replication and recombination machinery, and are particularly suited for extension by the telomerase reverse transcriptase. Helicases use the energy from NTP hydrolysis to track along DNA and disrupt base pairing. Here we review current findings concerning how helicases modulate several aspects of telomere form and function.
Figures
Similar articles
-
Telomere recombination and alternative telomere lengthening mechanisms.Front Biosci (Landmark Ed). 2013 Jan 1;18(1):1-20. doi: 10.2741/4084. Front Biosci (Landmark Ed). 2013. PMID: 23276906 Review.
-
Getting it done at the ends: Pif1 family DNA helicases and telomeres.DNA Repair (Amst). 2016 Aug;44:151-158. doi: 10.1016/j.dnarep.2016.05.021. Epub 2016 May 16. DNA Repair (Amst). 2016. PMID: 27233114 Free PMC article. Review.
-
Pif1p helicase, a catalytic inhibitor of telomerase in yeast.Science. 2000 Aug 4;289(5480):771-4. doi: 10.1126/science.289.5480.771. Science. 2000. PMID: 10926538
-
DNA helicases and their roles in cancer.DNA Repair (Amst). 2020 Dec;96:102994. doi: 10.1016/j.dnarep.2020.102994. Epub 2020 Oct 17. DNA Repair (Amst). 2020. PMID: 33137625 Review.
-
Resolving Roadblocks to Telomere Replication.Methods Mol Biol. 2019;1999:31-57. doi: 10.1007/978-1-4939-9500-4_2. Methods Mol Biol. 2019. PMID: 31127568 Review.
Cited by
-
Telomerase Regulation from Beginning to the End.Genes (Basel). 2016 Sep 14;7(9):64. doi: 10.3390/genes7090064. Genes (Basel). 2016. PMID: 27649246 Free PMC article. Review.
-
BMAL1 associates with chromosome ends to control rhythms in TERRA and telomeric heterochromatin.PLoS One. 2019 Oct 21;14(10):e0223803. doi: 10.1371/journal.pone.0223803. eCollection 2019. PLoS One. 2019. PMID: 31634367 Free PMC article.
-
Inactivation of the Sas2 histone acetyltransferase delays senescence driven by telomere dysfunction.EMBO J. 2010 Jan 6;29(1):158-70. doi: 10.1038/emboj.2009.314. Epub 2009 Oct 29. EMBO J. 2010. PMID: 19875981 Free PMC article.
-
Genome-wide comprehensive analysis of human helicases.Commun Integr Biol. 2011 Jan;4(1):118-37. doi: 10.4161/cib.4.1.13844. Commun Integr Biol. 2011. PMID: 21509200 Free PMC article.
-
Timeless preserves telomere length by promoting efficient DNA replication through human telomeres.Cell Cycle. 2012 Jun 15;11(12):2337-47. doi: 10.4161/cc.20810. Epub 2012 Jun 15. Cell Cycle. 2012. PMID: 22672906 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
